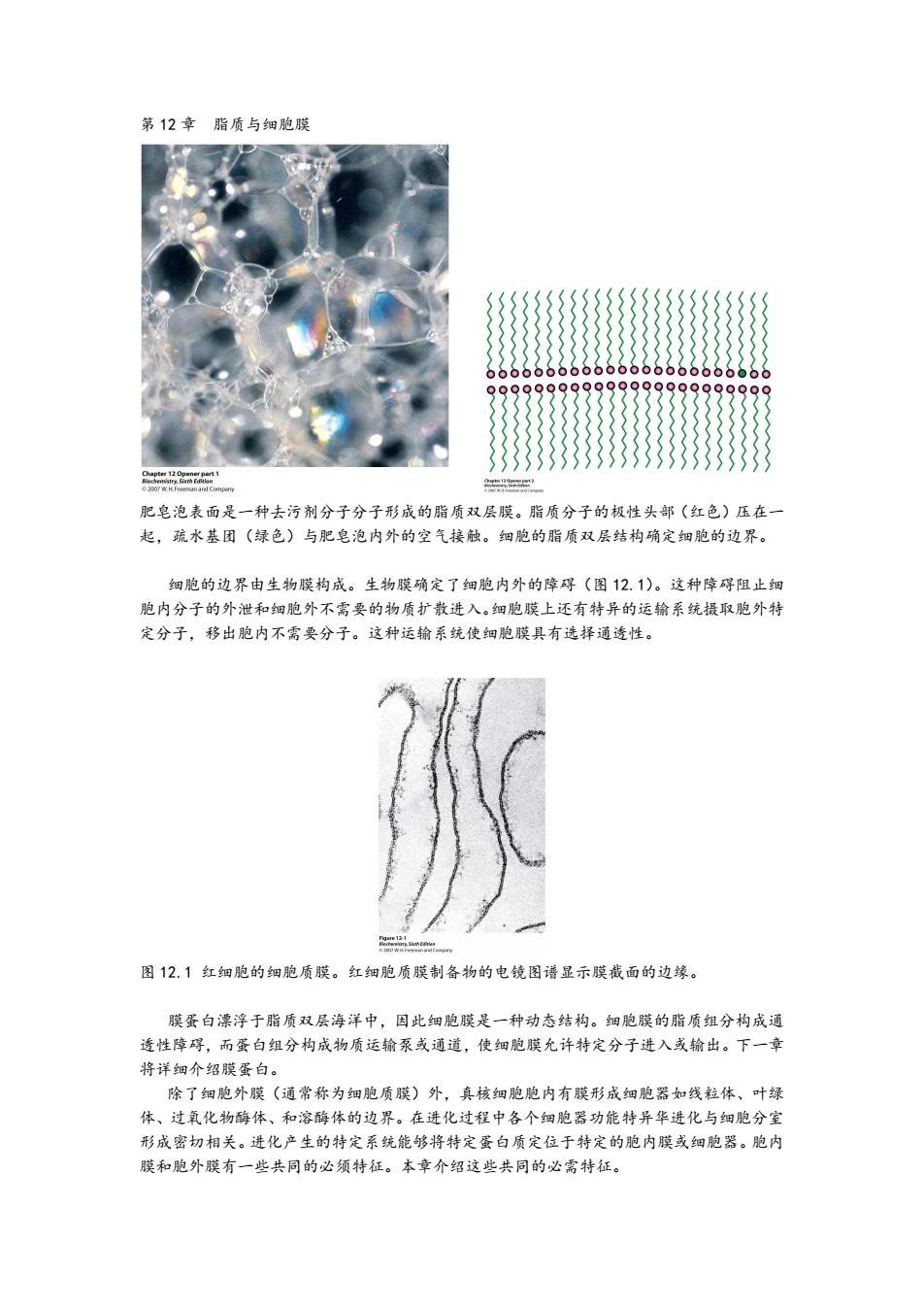
第12章脂质与细胞膜 肥皂泡表面是一种去污剂分子分子形成的脂质双层膜。脂质分子的极性头部(红色)压在一 起,疏水基团(绿色)与肥皂泡内外的空气接触。细胞的脂质双层结构确定细胞的边界。 细胞的边界由生物膜构成。生物膜确定了细胞内外的障碍(图12.1)。这种障碍阻止细 胞内分子的外泄和细胞外不需要的物质扩散进入。细胞膜上还有特异的运输系统摄取胞外特 定分子,移出胞内不需要分子。这种运输系统使细胞膜具有选择通透性。 图12.1红细胞的细胞质膜。红细胞质膜制备物的电镜图谱显示膜截面的边缘。 膜蛋白漂浮于脂质双层海洋中,因此细胞膜是一种动态结构。细胞膜的脂质组分构成通 透性障碍,而蛋白组分构成物质运输泵或通道,使细胞膜允许特定分子进入或输出。下一章 将详细介绍膜蛋白。 除了细胞外膜(通常称为细胞质膜)外,真核细胞胞内有膜形成细胞器如线粒体、叶绿 体、过氧化物酶体、和溶酶体的边界。在进化过程中各个细胞器功能特异华进化与细胞分室 形成密切相关。进化产生的特定系统能够将特定蛋白质定位于特定的胞内膜或细胞器。胞内 膜和胞外膜有一些共同的必须特征。本章介绍这些共同的必需特征
第 12 章 脂质与细胞膜 肥皂泡表面是一种去污剂分子分子形成的脂质双层膜。脂质分子的极性头部(红色)压在一 起,疏水基团(绿色)与肥皂泡内外的空气接触。细胞的脂质双层结构确定细胞的边界。 细胞的边界由生物膜构成。生物膜确定了细胞内外的障碍(图 12.1)。这种障碍阻止细 胞内分子的外泄和细胞外不需要的物质扩散进入。细胞膜上还有特异的运输系统摄取胞外特 定分子,移出胞内不需要分子。这种运输系统使细胞膜具有选择通透性。 图 12.1 红细胞的细胞质膜。红细胞质膜制备物的电镜图谱显示膜截面的边缘。 膜蛋白漂浮于脂质双层海洋中,因此细胞膜是一种动态结构。细胞膜的脂质组分构成通 透性障碍,而蛋白组分构成物质运输泵或通道,使细胞膜允许特定分子进入或输出。下一章 将详细介绍膜蛋白。 除了细胞外膜(通常称为细胞质膜)外,真核细胞胞内有膜形成细胞器如线粒体、叶绿 体、过氧化物酶体、和溶酶体的边界。在进化过程中各个细胞器功能特异华进化与细胞分室 形成密切相关。进化产生的特定系统能够将特定蛋白质定位于特定的胞内膜或细胞器。胞内 膜和胞外膜有一些共同的必须特征。本章介绍这些共同的必需特征

生物膜还有几种功能是生命必不可少的,如能量储存和信息转导。这些过程就是与生物 膜结合的蛋白质执行的。本章我们将介绍膜蛋白的普遍特性一一这些蛋白质如何存在于生物 膜的疏水环境?在这种疏水环境中如何执行自身功能?后续章节还要涉及这方面的内容。 结构和功能不同的细胞膜有一些共同的特征 虽然膜的结构和功能差异很大,但是它们有很多共同的特征: 1.膜是层类结构,只有两个分子厚度,形成内容物和外在环境之间封闭的边界。大多数生 物膜的厚度是60-100A(即6-10nm)。 2.膜的组分主要是脂质和蛋白质。脂质和蛋白质之间的质量比是1:4至4:1。膜含有碳 水化合物。这些糖分子与脂质或蛋白质连接。 3.膜的脂质分子小,有亲水和疏水两个部分。在水溶液环境中,这些脂质分子能够自动形 成封闭的水分子层,即脂质双层。这种双分子层是极性分子跨膜移动的障碍。 4.特殊蛋白质介导生物膜的不同功能。膜蛋白质可以充当泵、通道、受体、能量转导器、 和酶。脂质双层为处于脂质双层的酶蛋白分子发挥酶促作用提供了适宜的环境。 5.膜分子组装依靠非共价相互作用。膜蛋白和脂质分子之间有很多非共价作用。这些非共 价相互作用协同,将蛋白质和脂质分子结合在一起。 6.膜不对称。生物膜的两面总是不相同的。 7.生物膜是流动性结构。脂质分子沿脂质双层平面进行侧向水平移动,速度很快。膜蛋白 也能这样(除非蛋白质受特异的相互作用被锚定)。脂质分子和蛋白质不能跨膜旋转。 因此,生物膜可以看作是脂质和蛋白质之间有序排列的二维结构。 8. 大多数细胞膜两侧电极化,导致胞内呈电负性(通常是-60mV)。膜电位在物质运输、能 量转化、和激活等方面起关键作用(13章) 12.1脂肪酸是脂质分子的关键组分 脂质分子的疏水特性是脂质分子能够形成膜结构不可或缺的。大多数脂质分子的疏水特 性来自其脂肪酸组分的性质。 脂肪酸的命名基于母本碳氢化合物 脂肪酸是带有羧基的碳氢化合物。碳氢化合物的长度和饱和度有差异。脂肪酸的系统命 名法如下,将母本碳氢化合物的名称最后的字母e用oic acid替换即可。例如,C18饱和 脂肪酸称为octadecanoic acid(母本碳氢化合物名称是octadecane).。有一个双镀的CI8 脂肪酸叫octadecenoic acid,有两个双键的C18脂肪酸叫octadecadienoic acid,有三个 双键的C18脂肪酸叫octadecatrienoic acid。脂肪酸用数字命名的方法:18:0表示18碳 原子的脂肪酸,没有不饱和键:18:2表示18碳原子的脂肪酸,有两个双键。脂肪酸的羧基 发生解离,如软脂酸(16:0,pa1 mitic acid)和油酸(18:1)(图12.2)。 Palmitate (ionized form of palmitic acid) Oleate (ionized form of oleic acid) 图12.2两个脂肪酸的结构。软脂酸是含有16个碳原子的饱和脂肪酸。油酸是含有一个顺 式双键的18个碳原子脂肪酸
生物膜还有几种功能是生命必不可少的,如能量储存和信息转导。这些过程就是与生物 膜结合的蛋白质执行的。本章我们将介绍膜蛋白的普遍特性——这些蛋白质如何存在于生物 膜的疏水环境?在这种疏水环境中如何执行自身功能?后续章节还要涉及这方面的内容。 结构和功能不同的细胞膜有一些共同的特征 虽然膜的结构和功能差异很大,但是它们有很多共同的特征: 1. 膜是层类结构,只有两个分子厚度,形成内容物和外在环境之间封闭的边界。大多数生 物膜的厚度是 60 -100 A(即 6 -10 nm)。 2. 膜的组分主要是脂质和蛋白质。脂质和蛋白质之间的质量比是 1:4 至 4:1。膜含有碳 水化合物。这些糖分子与脂质或蛋白质连接。 3. 膜的脂质分子小,有亲水和疏水两个部分。在水溶液环境中,这些脂质分子能够自动形 成封闭的水分子层,即脂质双层。这种双分子层是极性分子跨膜移动的障碍。 4. 特殊蛋白质介导生物膜的不同功能。膜蛋白质可以充当泵、通道、受体、能量转导器、 和酶。脂质双层为处于脂质双层的酶蛋白分子发挥酶促作用提供了适宜的环境。 5. 膜分子组装依靠非共价相互作用。膜蛋白和脂质分子之间有很多非共价作用。这些非共 价相互作用协同,将蛋白质和脂质分子结合在一起。 6. 膜不对称。生物膜的两面总是不相同的。 7. 生物膜是流动性结构。脂质分子沿脂质双层平面进行侧向水平移动,速度很快。膜蛋白 也能这样(除非蛋白质受特异的相互作用被锚定)。脂质分子和蛋白质不能跨膜旋转。 因此,生物膜可以看作是脂质和蛋白质之间有序排列的二维结构。 8. 大多数细胞膜两侧电极化,导致胞内呈电负性(通常是-60mV)。膜电位在物质运输、能 量转化、和激活等方面起关键作用(13 章) 12.1 脂肪酸是脂质分子的关键组分 脂质分子的疏水特性是脂质分子能够形成膜结构不可或缺的。大多数脂质分子的疏水特 性来自其脂肪酸组分的性质。 脂肪酸的命名基于母本碳氢化合物 脂肪酸是带有羧基的碳氢化合物。碳氢化合物的长度和饱和度有差异。脂肪酸的系统命 名法如下,将母本碳氢化合物的名称最后的字母 e 用 oic acid 替换即可。例如,C18 饱和 脂肪酸称为 octadecanoic acid(母本碳氢化合物名称是 octadecane)。有一个双键的 C18 脂肪酸叫 octadecenoic acid,有两个双键的 C18 脂肪酸叫 octadecadienoic acid,有三个 双键的 C18 脂肪酸叫 octadecatrienoic acid。脂肪酸用数字命名的方法:18:0 表示 18 碳 原子的脂肪酸,没有不饱和键;18:2 表示 18 碳原子的脂肪酸,有两个双键。脂肪酸的羧基 发生解离,如软脂酸(16:0,palmitic acid)和油酸(18:1)(图 12.2)。 图 12.2 两个脂肪酸的结构。软脂酸是含有 16 个碳原子的饱和脂肪酸。油酸是含有一个顺 式双键的 18 个碳原子脂肪酸

脂肪酸碳原子的编号从羧基碳原子开始。第2号和第3号碳原子常被分别称为和β碳 原子。脂肪链最远的碳原子称为⊙碳原子(倒数第二位的碳原子称为⊙-2碳原子)。双键用△ 表示。双镀的位置标在△的右上角。而双键的构型在△前面用cis-或trans-表示。例如, cis-△’表示第9号碳原子和第10号碳原子之间有一个顺式双键:trans--△2表示在第2号 碳原子和第3号碳原子之间有一个反式双键。此外,也可以用倒数的形式(即▣-)表示双键。 例如o-3脂肪酸。在生理pH条件下,脂肪酸都是解离的,因此通常称为脂酸根,如palmitate 或hexadecanoate 脂肪酸之间的差异表现在长度和饱和度 生物体内的脂肪酸通常是偶数碳原子,典型的脂肪酸碳链长度在14至24碳原子之间(表 12.1)。最常见的脂肪酸碳原子长度是16碳原子和18碳原子脂肪酸。偶数碳原子脂肪酸占 优势与脂肪酸生物合成方式一致(参阅26章)。动物脂肪酸链几乎总是直链,没有分支。脂 肪链可能是饱和的,也可能含有双键。大多数不饱和脂肪酸的双键都是顺式(cⅰs)。脂肪链 含有多个双键的脂肪酸的双键之间至少间隔一个甲基。 TABLE 12.I Some naturally occurring fatty acids in animals Number of Number of Common carbons double bonds name Systematic name Formula 12 0 Laurate n-Dodecanoate CH3(CH,)COO 14 0 Myristate n-Tetradecanoate CH3(CH2)12COO- 0 Palmitate n-Hexadecanoate CH(CH,),COO 18 Stearate n-Octadecanoate CH.(CH,).COO 0 0 Arachidate n-Eicosanoate CH3(CH2)1sCOO 22 0 Behenate n-Docosanoate CH,(CH,)COO 0 Lignocerate n-Tetracosanoate CH;(CH,),COO 16 Palmitoleate c-△9-Hexadecenoate CH (CH,).CH-CH(CH,).COO Oleate cis-A-Octadecenoate CH,(CH,)CH-CH(CH,).COO 18 2 Linoleate cs,cis-A,△12. CH3 (CH)(CH-CHCH)(CH).COO Octadecadienoate 18 9 Linolenate all-ci-△9,A12,△15. CH CH,(CH-CHCH,)(CH,).COO Octadecatrienoate 20 4 Arachidonate all-cis△5,△8,△11,-A14 CH(CH,)(CH-CHCH)(CH.).COO Eicosatetraenoate Sixth Edition 2007W.H.Freeman and Company 脂质分子的脂肪酸性质主要取决于脂肪链长度和饱和度。不饱和脂肪酸的熔点比同等长 度的饱和脂肪酸低。例如,硬脂酸的熔点是69.6℃,而油酸(顺-9-十八碳单烯酸,oleic acid) 的熔点是13.4℃。含有多个双键的十八碳脂肪酸熔点更低。脂肪链的长度也影响脂肪链的 熔点。十六碳原子的饱和脂肪酸比十八碳原子的饱和脂肪酸低6.5度。因此脂肪链长度短、 不饱和度高能促进脂肪酸及其衍生物的流动性。 12.2有三类膜脂质 根据定义,脂质是水不溶、而在有机溶剂如氯仿中溶解的生物物质。脂质分子有不同的 生物功能:可以充当燃料、储存能量、信号传导途径的信号分子、和膜组分。后面章节将介 绍脂质分子前面的几个功能。此处我们集中讨论作为膜组分的脂质分子。膜脂质分子主要有 三类:磷脂、糖脂、和胆固醇。首先介绍真核生物和细菌的磷脂。虽然古生菌脂质的性质与 其它生物的膜脂质有很多共性,但是古生菌的脂质分子很独特
脂肪酸碳原子的编号从羧基碳原子开始。第 2 号和第 3 号碳原子常被分别称为和碳 原子。脂肪链最远的碳原子称为碳原子(倒数第二位的碳原子称为碳原子)。双键用△ 表示。双键的位置标在△的右上角。而双键的构型在△前面用 cis-或 trans-表示。例如, cis-△ 9表示第 9 号碳原子和第 10 号碳原子之间有一个顺式双键;trans-△ 2表示在第 2 号 碳原子和第 3 号碳原子之间有一个反式双键。此外,也可以用倒数的形式(即表示双键。 例如脂肪酸。在生理 pH 条件下,脂肪酸都是解离的,因此通常称为脂酸根,如 palmitate 或 hexadecanoate。 脂肪酸之间的差异表现在长度和饱和度 生物体内的脂肪酸通常是偶数碳原子,典型的脂肪酸碳链长度在14至24碳原子之间(表 12.1)。最常见的脂肪酸碳原子长度是 16 碳原子和 18 碳原子脂肪酸。偶数碳原子脂肪酸占 优势与脂肪酸生物合成方式一致(参阅 26 章)。动物脂肪酸链几乎总是直链,没有分支。脂 肪链可能是饱和的,也可能含有双键。大多数不饱和脂肪酸的双键都是顺式(cis)。脂肪链 含有多个双键的脂肪酸的双键之间至少间隔一个甲基。 脂质分子的脂肪酸性质主要取决于脂肪链长度和饱和度。不饱和脂肪酸的熔点比同等长 度的饱和脂肪酸低。例如,硬脂酸的熔点是 69.6℃,而油酸(顺-9-十八碳单烯酸,oleic acid) 的熔点是 13.4℃。含有多个双键的十八碳脂肪酸熔点更低。脂肪链的长度也影响脂肪链的 熔点。十六碳原子的饱和脂肪酸比十八碳原子的饱和脂肪酸低 6.5 度。因此脂肪链长度短、 不饱和度高能促进脂肪酸及其衍生物的流动性。 12.2 有三类膜脂质 根据定义,脂质是水不溶、而在有机溶剂如氯仿中溶解的生物物质。脂质分子有不同的 生物功能:可以充当燃料、储存能量、信号传导途径的信号分子、和膜组分。后面章节将介 绍脂质分子前面的几个功能。此处我们集中讨论作为膜组分的脂质分子。膜脂质分子主要有 三类:磷脂、糖脂、和胆固醇。首先介绍真核生物和细菌的磷脂。虽然古生菌脂质的性质与 其它生物的膜脂质有很多共性,但是古生菌的脂质分子很独特
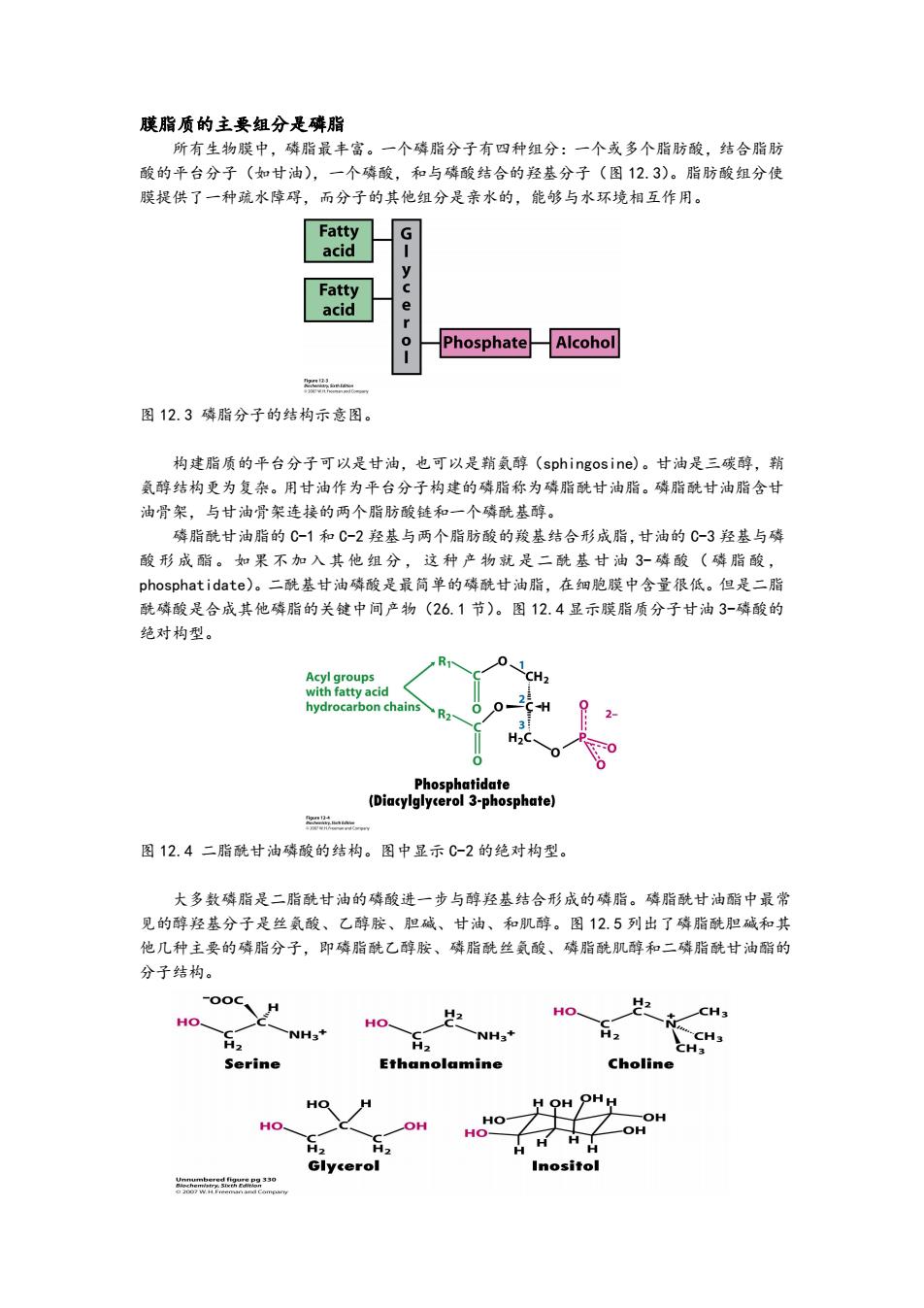
膜脂质的主要组分是磷脂 所有生物膜中,磷脂最丰富。一个磷脂分子有四种组分:一个或多个脂肪酸,结合脂肪 酸的平台分子(如甘油),一个磷酸,和与磷酸结合的羟基分子(图12.3)。脂肪酸组分使 膜提供了一种疏水障碍,而分子的其他组分是亲水的,能够与水环境相互作用。 Fatty G acid y Fatty acid e 0 Phosphate Alcohol 图12.3磷脂分子的结构示意图。 构建脂质的平台分子可以是甘油,也可以是鞘氨醇(sphingosine)。甘油是三碳醇,鞘 氨醇结构更为复杂。用甘油作为平台分子构建的磷脂称为磷脂酰甘油脂。磷脂酰甘油脂含甘 油骨架,与甘油骨架连接的两个脂肪酸链和一个磷酰基醇。 磷脂酰甘油脂的C1和C-2羟基与两个脂肪酸的羧基结合形成脂,甘油的C3羟基与磷 酸形成酯。如果不加入其他组分,这种产物就是二酰基甘油3-磷酸(磷脂酸, phosphatidate)。二酰基甘油磷酸是最简单的磷酰甘油脂,在细胞膜中含量很低。但是二脂 酰磷酸是合成其他磷脂的关键中间产物(26.1节)。图12.4显示膜脂质分子甘油3-磷酸的 绝对构型。 Acyl groups with fatty acid hydrocarbon chains H2 Phosphatidate (Diacylglycerol 3-phosphate) m 图12.4二脂酰甘油磷酸的结构。图中显示C-2的绝对构型。 大多数磷脂是二脂酰甘油的磷酸进一步与醇羟基结合形成的磷脂。磷脂酰甘油酯中最常 见的醇羟基分子是丝氨酸、乙醇胺、胆碱、甘油、和肌醇。图12.5列出了磷脂酰胆碱和其 他几种主要的磷脂分子,即磷脂酰乙醇胺、磷脂酰丝氨酸、磷脂酰肌醇和二磷脂酰甘油酯的 分子结构。 H H2 以2 HO CH3 HO HO NH3+ NH3+ CH3 H2 H2 CH3 Serine Ethanolamine Choline HO OH OH HO、 OH HO H2 H2 H Glycerol Inositol 30
膜脂质的主要组分是磷脂 所有生物膜中,磷脂最丰富。一个磷脂分子有四种组分:一个或多个脂肪酸,结合脂肪 酸的平台分子(如甘油),一个磷酸,和与磷酸结合的羟基分子(图 12.3)。脂肪酸组分使 膜提供了一种疏水障碍,而分子的其他组分是亲水的,能够与水环境相互作用。 图 12.3 磷脂分子的结构示意图。 构建脂质的平台分子可以是甘油,也可以是鞘氨醇(sphingosine)。甘油是三碳醇,鞘 氨醇结构更为复杂。用甘油作为平台分子构建的磷脂称为磷脂酰甘油脂。磷脂酰甘油脂含甘 油骨架,与甘油骨架连接的两个脂肪酸链和一个磷酰基醇。 磷脂酰甘油脂的 C-1 和 C-2 羟基与两个脂肪酸的羧基结合形成脂,甘油的 C-3 羟基与磷 酸 形成 酯。 如果 不 加入 其他 组分 ,这 种 产物 就是 二酰 基 甘油 3-磷 酸( 磷脂 酸, phosphatidate)。二酰基甘油磷酸是最简单的磷酰甘油脂,在细胞膜中含量很低。但是二脂 酰磷酸是合成其他磷脂的关键中间产物(26.1 节)。图 12.4 显示膜脂质分子甘油 3-磷酸的 绝对构型。 图 12.4 二脂酰甘油磷酸的结构。图中显示 C-2 的绝对构型。 大多数磷脂是二脂酰甘油的磷酸进一步与醇羟基结合形成的磷脂。磷脂酰甘油酯中最常 见的醇羟基分子是丝氨酸、乙醇胺、胆碱、甘油、和肌醇。图 12.5 列出了磷脂酰胆碱和其 他几种主要的磷脂分子,即磷脂酰乙醇胺、磷脂酰丝氨酸、磷脂酰肌醇和二磷脂酰甘油酯的 分子结构
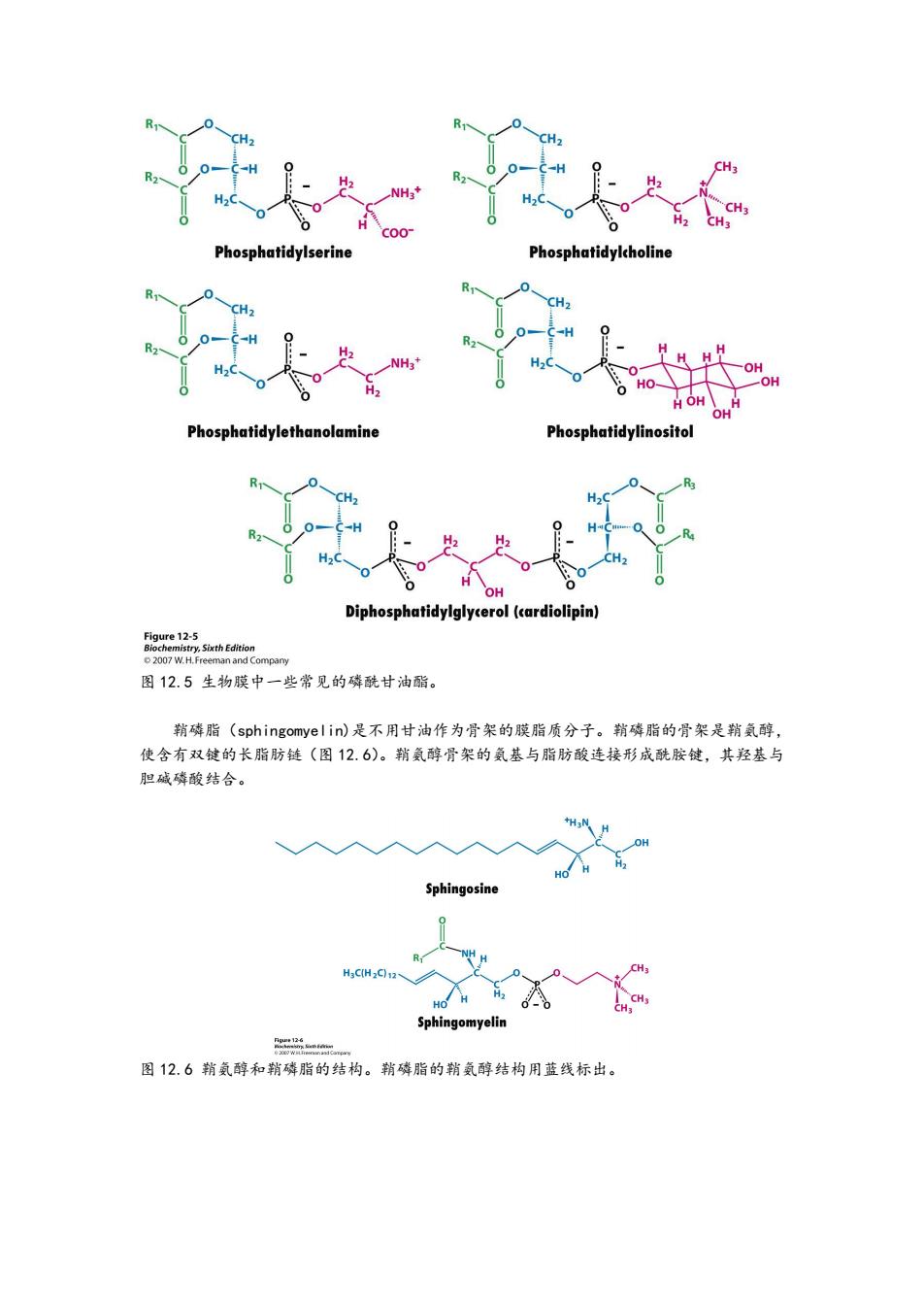
H NH CH C00 Phosphatidylserine Phosphatidylcholine H H OH -OH Phosphatidylethanolamine Phosphatidylinositol OH Diphosphatidylglycerol(cardiolipin) Figure 12-5 Biochemistry,Sixth Edition 2007 W.H.Freeman and Company 图12.5生物膜中一些常见的磷酰甘油酯。 鞘磷脂(sphingomye l in)是不用甘油作为骨架的膜脂质分子。鞘磷脂的骨架是鞘氨醇, 使含有双键的长脂肪链(图12.6)。鞘氨醇骨架的氨基与脂肪酸连接形成酰胺键,其羟基与 胆碱磷酸结合。 H OH HO Sphingosine H3CHzC92、 Sphingomyelin 二 图12.6鞘氨醇和鞘磷脂的结构。鞘磷脂的鞘氨醇结构用蓝线标出
图 12.5 生物膜中一些常见的磷酰甘油酯。 鞘磷脂(sphingomyelin)是不用甘油作为骨架的膜脂质分子。鞘磷脂的骨架是鞘氨醇, 使含有双键的长脂肪链(图 12.6)。鞘氨醇骨架的氨基与脂肪酸连接形成酰胺键,其羟基与 胆碱磷酸结合。 图 12.6 鞘氨醇和鞘磷脂的结构。鞘磷脂的鞘氨醇结构用蓝线标出