
2生化进化 地球大约有45亿年历史。有大量令人信服的化石证据显示35亿年前的生物形态(很 可能是生物化学过程)与一些现代细菌相似。定向研究和偶然发现所获得的结果能勾画出前 生命世界进化到现代生物世界的途径。虽然这种进化途径是一种假设,但看上去很有道理。 有些进化细节还不能完全肯定,尤其是最早发生的事件。然而,思考这条进化途径的中间步 骤和相关生化问题的答案能很好地理解现代生物的生化过程。这些进化联系使很多生物化学 内容易于理解。 我们可以将原始生命进化到现代生物的过程分成不同的阶段,但是读者应该记住实 际的进化过程不会像此处陈述的如此清晰。第一阶段非生命过程产生生命物质。采用非生命 过程产生第一代重要的生命分子一一核酸、蛋白质、碳水化合物和脂肪。第一阶段形成活细 胞。随着时间的推移,这些系统越来越复杂抑制能够形成活细胞。第三阶段进化出一种机制, 能够将化学能和太阳能转化成生化过程可以利用的能量。这些能量转换过程与核酸、蛋白质 和其它关键物质单体合成途径关联。随着能量转化过程和生物合成途径的建立,就出现了不 同的单细胞生物。第四阶段建立一种机制使细胞针对不同的环境改变其生物化学过程,甚至 发展成多细胞生物。 本章介绍生命进化过程所面临的关键问题。后面的章节讲究这些关键问题进行阐述。 探讨这些生化过程的可能起源是进化的用途更容易理解。 自然选择是进化的关键动力之一,创造了一种生态小环境适于那些能够生化适应的物 种。左边是盐池,盐浓度高达1.5M以上,几乎不适合生物生存。但是有些halophilic archaea如Haloferax mediterranei(右边),能够适应这种恶劣环境。它们有相应的生 化过程适应这种环境。[(Left)KajR.Svensson/Science Photo Library/Photo Researchers;(right)Wanner/Eye of Science/Photo Researchers.] 2.1生命体所用的关键有机分子 地球形成约10亿年后,地球上出现生命。在生命出现之前,还需要发生另一个事件 一一利用环境中简单分子合成生命需要的关键的有机分子。核酸和蛋白质的组分是比较复杂 的有机分子,人们可能会认为只有先进的合成路线才能合成这些有机分子。但是,这中想法 是不符合实际的。那么,这些有机分子是如如何合成的呢? 2.1.1.生物大分子的很多组分可以用简单的、前生命反应制造 有几个理论阐述了前生命世界的环境条件,但是没有一种理论完美无缺。有一种理论认 为早期地球还原性强,富含甲烷、氨气、水和氢气,这种大气受到太阳的大剂量辐射和光照。 为了讨论起见,我们假定这些条件就是地球前生命时期的实际环境。在这样的环境条件下能 够产生复杂的有机分子吗?1950年代Stanley Miller和Harold Urey给出了答案。他们
2 生化进化 地球大约有 45 亿年历史。有大量令人信服的化石证据显示 35 亿年前的生物形态(很 可能是生物化学过程)与一些现代细菌相似。定向研究和偶然发现所获得的结果能勾画出前 生命世界进化到现代生物世界的途径。虽然这种进化途径是一种假设,但看上去很有道理。 有些进化细节还不能完全肯定,尤其是最早发生的事件。然而,思考这条进化途径的中间步 骤和相关生化问题的答案能很好地理解现代生物的生化过程。这些进化联系使很多生物化学 内容易于理解。 我们可以将原始生命进化到现代生物的过程分成不同的阶段,但是读者应该记住实 际的进化过程不会像此处陈述的如此清晰。第一阶段非生命过程产生生命物质。采用非生命 过程产生第一代重要的生命分子——核酸、蛋白质、碳水化合物和脂肪。第一阶段形成活细 胞。随着时间的推移,这些系统越来越复杂抑制能够形成活细胞。第三阶段进化出一种机制, 能够将化学能和太阳能转化成生化过程可以利用的能量。这些能量转换过程与核酸、蛋白质 和其它关键物质单体合成途径关联。随着能量转化过程和生物合成途径的建立,就出现了不 同的单细胞生物。第四阶段建立一种机制使细胞针对不同的环境改变其生物化学过程,甚至 发展成多细胞生物。 本章介绍生命进化过程所面临的关键问题。后面的章节讲究这些关键问题进行阐述。 探讨这些生化过程的可能起源是进化的用途更容易理解。 自然选择是进化的关键动力之一,创造了一种生态小环境适于那些能够生化适应的物 种。左边是盐池,盐浓度高达1.5 M以上,几乎不适合生物生存。但是有些halophilic archaea 如 Haloferax mediterranei (右边), 能够适应这种恶劣环境。它们有相应的生 化过程适应这种环境。[(Left) Kaj R. Svensson/Science Photo Library/Photo Researchers; (right) Wanner/Eye of Science/Photo Researchers.] 2.1 生命体所用的关键有机分子 地球形成约 10 亿年后,地球上出现生命。在生命出现之前,还需要发生另一个事件 ——利用环境中简单分子合成生命需要的关键的有机分子。核酸和蛋白质的组分是比较复杂 的有机分子,人们可能会认为只有先进的合成路线才能合成这些有机分子。但是,这中想法 是不符合实际的。那么,这些有机分子是如如何合成的呢? 2.1.1. 生物大分子的很多组分可以用简单的、前生命反应制造 有几个理论阐述了前生命世界的环境条件,但是没有一种理论完美无缺。有一种理论认 为早期地球还原性强,富含甲烷、氨气、水和氢气,这种大气受到太阳的大剂量辐射和光照。 为了讨论起见,我们假定这些条件就是地球前生命时期的实际环境。在这样的环境条件下能 够产生复杂的有机分子吗?1950 年代 Stanley Miller 和 Harold Urey 给出了答案。他们
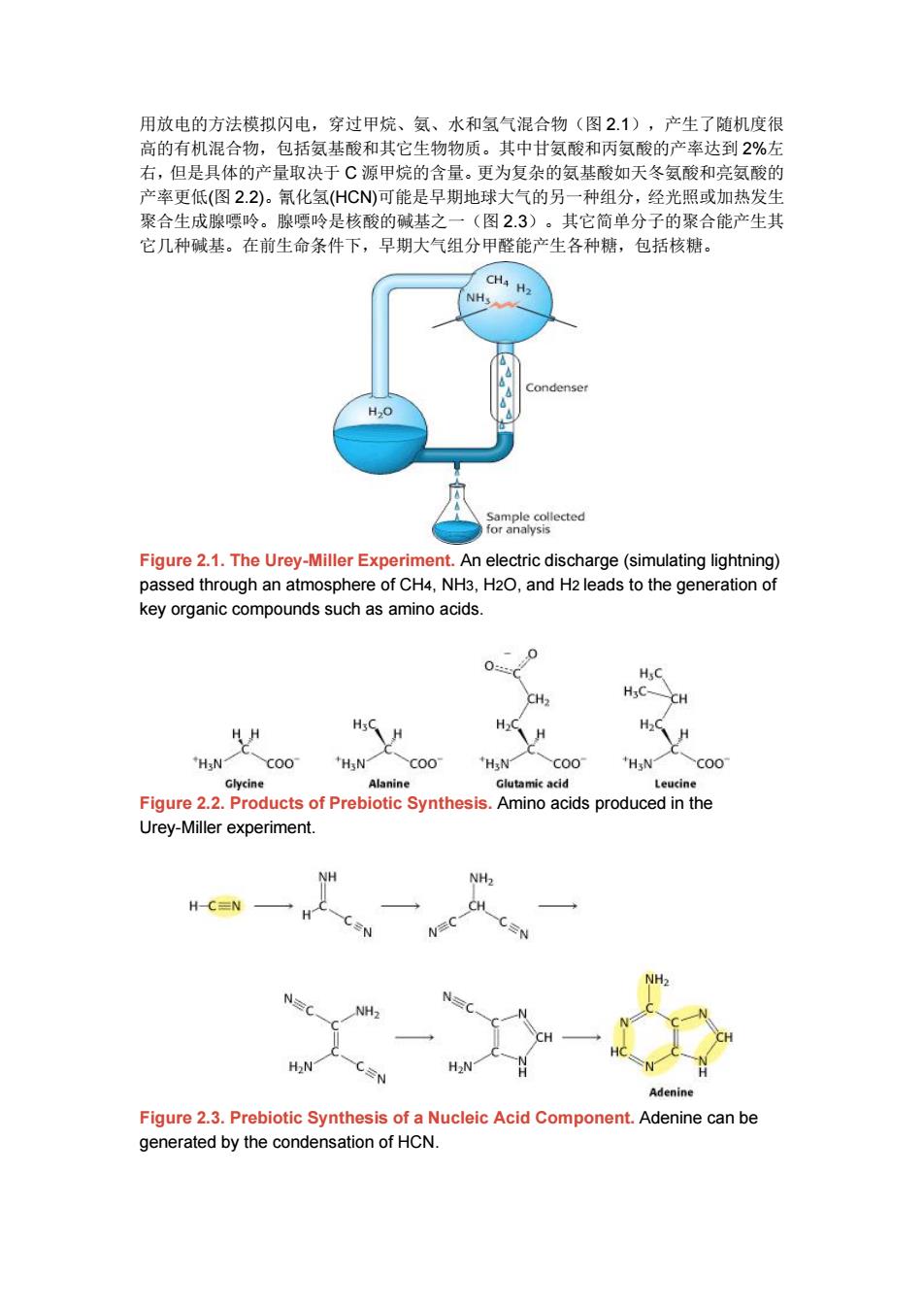
用放电的方法模拟闪电,穿过甲烷、氨、水和氢气混合物(图2.1),产生了随机度很 高的有机混合物,包括氨基酸和其它生物物质。其中甘氨酸和丙氨酸的产率达到2%左 右,但是具体的产量取决于C源甲烷的含量。更为复杂的氨基酸如天冬氨酸和亮氨酸的 产率更低(图22)。氰化氢(HCN)可能是早期地球大气的另一种组分,经光照或加热发生 聚合生成腺嘌呤。腺嘌呤是核酸的碱基之一(图2.3)。其它简单分子的聚合能产生其 它几种碱基。在前生命条件下,早期大气组分甲醛能产生各种糖,包括核糖。 CH4 H2 Condenser H20 Sample collected for analysis Figure 2.1.The Urey-Miller Experiment.An electric discharge(simulating lightning) passed through an atmosphere of CH4,NH3,H2O,and H2 leads to the generation of key organic compounds such as amino acids. 0 0 HyC CH2 CH HsC H HH H H H C00 HsN COO "HsN COO Glycine Alanine Glutamic acid Leucine Figure 2.2.Products of Prebiotic Synthesis.Amino acids produced in the Urey-Miller experiment. NH NH H-CEN H NH2 N NH> H2NT CN Adenine Figure 2.3.Prebiotic Synthesis of a Nucleic Acid Component.Adenine can be generated by the condensation of HCN
用放电的方法模拟闪电,穿过甲烷、氨、水和氢气混合物(图 2.1),产生了随机度很 高的有机混合物,包括氨基酸和其它生物物质。其中甘氨酸和丙氨酸的产率达到 2%左 右,但是具体的产量取决于 C 源甲烷的含量。更为复杂的氨基酸如天冬氨酸和亮氨酸的 产率更低(图 2.2)。氰化氢(HCN)可能是早期地球大气的另一种组分,经光照或加热发生 聚合生成腺嘌呤。腺嘌呤是核酸的碱基之一(图 2.3)。其它简单分子的聚合能产生其 它几种碱基。在前生命条件下,早期大气组分甲醛能产生各种糖,包括核糖。 Figure 2.1. The Urey-Miller Experiment. An electric discharge (simulating lightning) passed through an atmosphere of CH4, NH3, H2O, and H2 leads to the generation of key organic compounds such as amino acids. Figure 2.2. Products of Prebiotic Synthesis. Amino acids produced in the Urey-Miller experiment. Figure 2.3. Prebiotic Synthesis of a Nucleic Acid Component. Adenine can be generated by the condensation of HCN

2.2进化需要复制、变异、和压力 有了所需要的单体后,如何制造生命系统并进行进化呢?在生命出现之前,简单分子系 统要进化成具有生命特征的复杂化学系统。为了弄清楚进化是如何发生的,我们要研究进化 的过程。如果它们是分子的简单集合或它们是相互竞争的生物群体,则进化系统几个共同的 原则。(1)能够进化的系统必须具备繁殖或复制的能力。如果没有复制能力,这个种生物或 分子就会灭绝。例如,核糖核酸会降解。能够复制的分子即使每个分子个体的寿命短暂,但 是由于它能够复制因此依然存在于种群中。(2)能够进化的系统必须具备变异的能力,使复 制的产品发生变化。如果复制极度完美,产品与亲本一模一样,就无法发生进化。在2.2.5 节我们将讨论生命系统变异的本质。(3)能够进化的系统内部各成员处于竞争状态。这种竞 争就是自然选择,其结果是进化。变异产生了不同分子群体。有些变异分子对环境的适应力 更优越,这样的变异份子复制能力比亲本分子还强,它们在群体中的相对比例会大幅提高。 变异石发生在分子水平上,但是变异导致的生存优势却显示在生物个体水平上。 2.2.1.在体外能够证实进化原则 进化发生在分子水平上的证据是什么?1967年Sol Spiegelman指出复制分子在实验 过程中能够进化成新的分子,这样的实验使他能够在试管中观察分子进化。噬菌体Q邱 的基因组是单链RNA分子,长度有3300碱基,其复制需要QB复制酶。Spiegelman 将Qβ复制酶与Qβ混合,在不限时也无其它选择压力的情况下,系统能够将前体核苷酸 转化成QB RNA。子代分子的组成与亲本没有差异。如果施加选择压力,子代分子的组 成与亲本的差异就非常大。例如,将复制时间从20分钟缩短成5分钟,复制75代后, 子代明显优势分子的长度是550碱基。这种子代分子复制的速度是QB RNA亲本分子 的15倍(图2.4)。Spiegelman采用其它的选择压力,如限制前体的浓度或添加抑制 剂,就会出现能够适应新环境的具有复制优势的新分子。 QB 3300 bases 550 bases 25 50 Generation number Figure2.4.在试管中的进化.施加选择性条件,Q B RNA快速复制产生的RNA种类。 绿色和蓝色曲线分别代表复制过程中先积累后消失的中等大小的RNA分子。 这些研究证实进化过程依赖含有QB复制酶的RNA复制机器。如同第1章所谈到的, 核酸分子最巧妙的特征之一是分子的复制机制遵循分子的结构原则。这些观察提示核酸 能够进行自我复制。实际上的研究结果显示单链核酸能够作为模板复制互补的另一条单 链核酸,这种合成是自主发生的、无需生物合成机器驱动。但是研究人员至今还没有发 现用简单其实材料进行RNA分子自我复制的条件。 2.2.2.RNA能充当催化剂 要有超越简单复制的能力就得有特殊的催化剂。催化剂是能够加速特定化学反应,但本 身不会发生化学变化的分子。在第8章和第9章我们将详细讨论催化剂的性质。有些催化剂 的特异性很高,它们仅促进一些化学反应但不影响与这些反应相关性很高的另一些化学反
2.2 进化需要复制、变异、和压力 有了所需要的单体后,如何制造生命系统并进行进化呢?在生命出现之前,简单分子系 统要进化成具有生命特征的复杂化学系统。为了弄清楚进化是如何发生的,我们要研究进化 的过程。如果它们是分子的简单集合或它们是相互竞争的生物群体,则进化系统几个共同的 原则。(1)能够进化的系统必须具备繁殖或复制的能力。如果没有复制能力,这个种生物或 分子就会灭绝。例如,核糖核酸会降解。能够复制的分子即使每个分子个体的寿命短暂,但 是由于它能够复制因此依然存在于种群中。(2)能够进化的系统必须具备变异的能力,使复 制的产品发生变化。如果复制极度完美,产品与亲本一模一样,就无法发生进化。在 2.2.5 节我们将讨论生命系统变异的本质。(3)能够进化的系统内部各成员处于竞争状态。这种竞 争就是自然选择,其结果是进化。变异产生了不同分子群体。有些变异分子对环境的适应力 更优越,这样的变异份子复制能力比亲本分子还强,它们在群体中的相对比例会大幅提高。 变异石发生在分子水平上,但是变异导致的生存优势却显示在生物个体水平上。 2.2.1. 在体外能够证实进化原则 进化发生在分子水平上的证据是什么?1967 年 Sol Spiegelman 指出复制分子在实验 过程中能够进化成新的分子,这样的实验使他能够在试管中观察分子进化。噬菌体 Q 的基因组是单链 RNA 分子,长度有 3300 碱基,其复制需要 Q复制酶。Spiegelman 将 Q复制酶与 Q混合,在不限时也无其它选择压力的情况下,系统能够将前体核苷酸 转化成 Q RNA。子代分子的组成与亲本没有差异。如果施加选择压力,子代分子的组 成与亲本的差异就非常大。例如,将复制时间从 20 分钟缩短成 5 分钟,复制 75 代后, 子代明显优势分子的长度是 550 碱基。这种子代分子复制的速度是 Q RNA 亲本分子 的 15 倍(图 2.4)。Spiegelman 采用其它的选择压力,如限制前体的浓度或添加抑制 剂,就会出现能够适应新环境的具有复制优势的新分子。 Figure 2.4. 在试管中的进化. 施加选择性条件,Q RNA快速复制产生的RNA种类。 绿色和蓝色曲线分别代表复制过程中先积累后消失的中等大小的RNA分子。 这些研究证实进化过程依赖含有Q复制酶的RNA复制机器。如同第1章所谈到的, 核酸分子最巧妙的特征之一是分子的复制机制遵循分子的结构原则。这些观察提示核酸 能够进行自我复制。实际上的研究结果显示单链核酸能够作为模板复制互补的另一条单 链核酸,这种合成是自主发生的、无需生物合成机器驱动。但是研究人员至今还没有发 现用简单其实材料进行RNA分子自我复制的条件。 2.2.2. RNA 能充当催化剂 要有超越简单复制的能力就得有特殊的催化剂。催化剂是能够加速特定化学反应,但本 身不会发生化学变化的分子。在第 8 章和第 9 章我们将详细讨论催化剂的性质。有些催化剂 的特异性很高,它们仅促进一些化学反应但不影响与这些反应相关性很高的另一些化学反

应。这样的催化剂促使途径的化学反应进行,但不催化另一些代谢途径的发生。直到1980 年代,我们才认识到生物催化剂不仅只局限于蛋白质。Tom Cech和Sidney Altman独立 地发现了一些RNA分子能够充当生物催化剂。这些RNA分子充当的催化剂叫 ribozyme。Ribozyme的发现提示生命进化过程中催化RNA分子起重要作用。 RNA分子的催化活性与RNA分子结构特异、复杂密切相关。垂头状核酶 (hammerhead ribozyme)就是一个例子。RNA垂头状结构首先在植物病毒中发现(图 2.5)。这种RNA分子能促进RNA分子在特定位点发生裂解。对一些病毒生命周期而 言,这种裂解是必需的。核酶催化的RNA裂解需要Mg2+离子或其他离子。核酶与底物 RNA分子形成复合物后呈反应活化状态。 ②在本书中出现的此图标表示生物化学网站w.whfreeman.com/biochem5有一些 资源供读者搜索。该图标表示你在生物化学网站能观察三维立体图谱。登陆网站、选择 章节和数字编号。 (®) (A) Ribozyme GUAGU A Cleavage GCCGG site AAGGCCG Cleavage CCAGCCG-5 site GGUCGGC-3 A C A Substrate G G UA 35 Figure 2.5.Catalytic RNA.(A)The base-pairing pattern of a"hammerhead" ribozyme and its substrate.(B)The folded conformation of the complex.The ribozyme cleaves the bond at the cleavage site.The paths of the nucleic acid backbones are highlighted in red and blue. RNA分子具有特异性结合和催化的性能。这类分子的存在使早期生命形式表现为 “RNA世界”成为可行。在RNA世界,所有主要作用(包括信息的存储、生命体遗传、 生物合成和能量代谢的催化等)都依赖于生命体内RNA分子。 2.2.3.氨基酸及其聚合物起生物合成和催化作用 在早期的RNA世界,复制RNA分子数量的增加将耗尽前生命时期长期积累的RNA合 成单体。RNA合成单体的缺乏有助于它们更换合成机制。有很多可能的途径。当初产生 的那些合成途径生存下来,就成为现代生物体所采用的生物合成途径。一个明显的事实 是氨基酸成为RNA碱基的合成原料(图2.6)一种氨基酸作为核心,碱基的其余部分添 加到这个核心上,合成核酸的嘌呤碱基(腺嘌呤和鸟嘌呤)和嘧啶碱基(尿嘧啶和胞嘧啶)。 此外,碱基的氮原子来自天门冬氨酸的氨基和谷氨酰胺侧链氨基
应。这样的催化剂促使途径的化学反应进行,但不催化另一些代谢途径的发生。直到 1980 年代,我们才认识到生物催化剂不仅只局限于蛋白质。Tom Cech 和 Sidney Altman 独立 地发现了一些 RNA 分子能够充当生物催化剂。这些 RNA 分子充当的催化剂叫 ribozyme。Ribozyme 的发现提示生命进化过程中催化 RNA 分子起重要作用。 RNA 分子的催化活性与 RNA 分子结构特异、复杂密切相关。垂头状核酶 (hammerhead ribozyme)就是一个例子。RNA 垂头状结构首先在植物病毒中发现(图 2.5)。这种 RNA 分子能促进 RNA 分子在特定位点发生裂解。对一些病毒生命周期而 言,这种裂解是必需的。核酶催化的 RNA 裂解需要 Mg2+离子或其他离子。核酶与底物 RNA 分子形成复合物后呈反应活化状态。 在本书中出现的此图标表示生物化学网站 www.whfreeman.com/biochem5 有一些 资源供读者搜索。该图标表示你在生物化学网站能观察三维立体图谱。登陆网站、选择 章节和数字编号。 Figure 2.5. Catalytic RNA. (A) The base-pairing pattern of a "hammerhead" ribozyme and its substrate. (B) The folded conformation of the complex. The ribozyme cleaves the bond at the cleavage site. The paths of the nucleic acid backbones are highlighted in red and blue. RNA分子具有特异性结合和催化的性能。这类分子的存在使早期生命形式表现为 “RNA世界”成为可行。在RNA世界,所有主要作用(包括信息的存储、生命体遗传、 生物合成和能量代谢的催化等)都依赖于生命体内RNA分子。 2.2.3. 氨基酸及其聚合物起生物合成和催化作用 在早期的RNA世界,复制RNA分子数量的增加将耗尽前生命时期长期积累的RNA合 成单体。RNA合成单体的缺乏有助于它们更换合成机制。有很多可能的途径。当初产生 的那些合成途径生存下来,就成为现代生物体所采用的生物合成途径。一个明显的事实 是氨基酸成为RNA碱基的合成原料(图2.6)一种氨基酸作为核心,碱基的其余部分添 加到这个核心上,合成核酸的嘌呤碱基(腺嘌呤和鸟嘌呤)和嘧啶碱基(尿嘧啶和胞嘧啶)。 此外,碱基的氮原子来自天门冬氨酸的氨基和谷氨酰胺侧链氨基

NH;+ OOC-CH2 H Glycine C00 00c NH, Aspartic acid C00 H2C H Purine Aspartic acid Pyrimidine H:N Glutamine Figure2.6.RNA碱基的生物合成。氨基酸是嘌呤碱基和嘧啶碱基生物合成的原料。 氨基酸的化学样式比核酸多些,因为氨基酸侧链的化学差异性更大。氨基酸之间的 化学连接叫肽键,用肽键将氨基酸聚合起来的化合物叫多肽(图2.7)。多肽能够执行 核酶的功能。多肽链自动折叠形成的三维结构由多肽的氨基酸序列决定。这种三维结构 使之能够与其他分子的相互作用具有高度特异性,有助于蛋白质在生命进化过程中执行 更多的功能,使之成为现代生物体的优势分子。今天大多数生物催化剂(酶)不再是核 酸,而是蛋白质。 Peptide bonds Amino Amino Amino acid 1 acid 2 acid 3 Figure2.7.另一种具有功能的聚合物。蛋白质是用肽键连接的氨基酸聚合物。 2.2.4.指导多肽合成的RNA模板将RNA世界与蛋白质世界关联在一起 由于多肽的结构不能像核酸那样进行自我复制,因此进化早期多肽的作用有限。然 而多肽可以被间接包含在进化过程中。例如,若一个特定的多肽有助于某类RNA的复制 和生存,这类RNA分子将进化成一种核酶来促进这样的多肽合成。用这种方法制造多肽 有几个限制。(1)这种方式好像只能制造相对较短的多肽。(2)很难精确地复制出原 来序列的多肽链。(3)不同的核酶需要不同的多肽链。如果RNA分子的碱基序列能直 接指导多肽的氨基酸序列合成时,进化就达到了一个关键节点。密码子出现建立了RNA 三碱基序列与氨基酸之间的特异联系。我们现在将这套三碱基序列成为遗传密码。对遗 传信息,即RNA模板,进行解码叫翻译。进行翻译的场所是核糖体,还有一些相关因子 参与。多肽合成的模式见图2.8
Figure 2.6. RNA碱基的生物合成。氨基酸是嘌呤碱基和嘧啶碱基生物合成的原料。 氨基酸的化学样式比核酸多些,因为氨基酸侧链的化学差异性更大。氨基酸之间的 化学连接叫肽键,用肽键将氨基酸聚合起来的化合物叫多肽(图2.7)。多肽能够执行 核酶的功能。多肽链自动折叠形成的三维结构由多肽的氨基酸序列决定。这种三维结构 使之能够与其他分子的相互作用具有高度特异性,有助于蛋白质在生命进化过程中执行 更多的功能,使之成为现代生物体的优势分子。今天大多数生物催化剂(酶)不再是核 酸,而是蛋白质。 Figure 2.7. 另一种具有功能的聚合物。蛋白质是用肽键连接的氨基酸聚合物。 2.2.4. 指导多肽合成的RNA模板将RNA世界与蛋白质世界关联在一起 由于多肽的结构不能像核酸那样进行自我复制,因此进化早期多肽的作用有限。然 而多肽可以被间接包含在进化过程中。例如,若一个特定的多肽有助于某类RNA的复制 和生存,这类RNA分子将进化成一种核酶来促进这样的多肽合成。用这种方法制造多肽 有几个限制。(1)这种方式好像只能制造相对较短的多肽。(2)很难精确地复制出原 来序列的多肽链。(3)不同的核酶需要不同的多肽链。如果RNA分子的碱基序列能直 接指导多肽的氨基酸序列合成时,进化就达到了一个关键节点。密码子出现建立了RNA 三碱基序列与氨基酸之间的特异联系。我们现在将这套三碱基序列成为遗传密码。对遗 传信息,即RNA模板,进行解码叫翻译。进行翻译的场所是核糖体,还有一些相关因子 参与。多肽合成的模式见图2.8