
第十三章膜通道和泵 m7r气rm- 3 经过单一膜通道的离子流动(在左边的示意图中用红色表示通道)可以用全自动膜片钳测定。 这种技术能够记录通道开放和关闭状态的电流。 生物膜的脂质双层结构本身是离子和极性分子的通透障碍。但是要维持正常的细胞功 能,生物膜必须允许一些离子和极性分子通过。两类膜蛋白,即泵和通道,使生物膜具有这 样的功能。泵的能量来自能源分子如ATP的水解,或者是光能。泵能利用这些能源驱动离 子或分子作热力学的逆向运输(主动运输)。相反,通道不用能量,只是允许离子或分子作 热力学顺向运输(被动运输)。 泵是一种能量传导装置,能够将一种形式的自由能转化成另一种形式的自由能。ATP驱 动的泵油两种,P型ATP酶和和含有ATP结合盒(ABC)的运输器(transporter)。这两种 泵与ATP结合、水解ATP导致泵分子构型转化,使泵结合的离子被跨膜运输。另一种机制 利用离子梯度驱动其它物质跨膜运输。大肠杆菌乳糖运输器是这类次级运输器的一个例子。 乳糖运输器负责细菌从环境摄取特定糖分子。细胞膜有很多这类运输器。这些运输器的表达 决定了细胞所摄取物质的种类。因此调节运输器的表达是控制细胞代谢的主要手段。 泵能够建立特定离子持久的跨膜梯度。特定的离子通道允许这些离子迅速跨膜运输到浓 度低的一面(被动运输)。由于这些离子通道允许一些离子跨膜流过,而另一些离子(甚至 是那些与可跨膜流过离子密切相关的离子)不能通过,因此这些通道成为生物化学最迷人的 分子。这些门控离子通道在执行神经系统功能方面起中心作用。神经系统充当导线,允许精 细切换神经信号的快速流动。 最后,我们讨论另一种种通道,即细胞与细胞之间的通道(或缝隙连接,gap-junction).。 这种通道允许离子或代谢物质在细胞之间进行运输。例如分析连接的细胞间物质运输负责心 脏跳动时肌肉细胞同步收缩。 运输器的表达在很大程度上确定了特定细胞的代谢活性 各种细胞表达一套独有的细胞质膜运输器。由于这些运输器在很大程度上确定了细胞从 环境摄取离子和代谢物的模式,因此细胞表达的运输器组合非常重要。有些情况下,一个细 胞所表达的运输器组合就决定了这个细胞的性质,因为这个细胞只能摄取特定底物从而只能 执行特定的生物化学反应。 葡萄糖代谢能用来解释这种观,点。如同我们在第16章葡萄糖代谢部分将详细讨论的, 组织之间的差异在于他们利用不同分子充当能源的能力。哪个组织能够利用葡萄糖主要取决 于它们表达葡萄糖运输器的情况。这些葡萄糖运输器包括GLUT1,GLUT2,GLUT3,GLUT4, 和GLUT5。例如GLUT3只在神经元和集中其它类型的细胞表达。在葡萄糖浓度相对较低 时,GLUT3与葡萄糖结合相对较紧。在控制和整合活体代谢方面,这些运输器的表达非常
第十三章 膜通道和泵 经过单一膜通道的离子流动(在左边的示意图中用红色表示通道)可以用全自动膜片钳测定。 这种技术能够记录通道开放和关闭状态的电流。 生物膜的脂质双层结构本身是离子和极性分子的通透障碍。但是要维持正常的细胞功 能,生物膜必须允许一些离子和极性分子通过。两类膜蛋白,即泵和通道,使生物膜具有这 样的功能。泵的能量来自能源分子如 ATP 的水解,或者是光能。泵能利用这些能源驱动离 子或分子作热力学的逆向运输(主动运输)。相反,通道不用能量,只是允许离子或分子作 热力学顺向运输(被动运输)。 泵是一种能量传导装置,能够将一种形式的自由能转化成另一种形式的自由能。ATP 驱 动的泵油两种,P-型 ATP 酶和和含有 ATP 结合盒(ABC)的运输器(transporter)。这两种 泵与 ATP 结合、水解 ATP 导致泵分子构型转化,使泵结合的离子被跨膜运输。另一种机制 利用离子梯度驱动其它物质跨膜运输。大肠杆菌乳糖运输器是这类次级运输器的一个例子。 乳糖运输器负责细菌从环境摄取特定糖分子。细胞膜有很多这类运输器。这些运输器的表达 决定了细胞所摄取物质的种类。因此调节运输器的表达是控制细胞代谢的主要手段。 泵能够建立特定离子持久的跨膜梯度。特定的离子通道允许这些离子迅速跨膜运输到浓 度低的一面(被动运输)。由于这些离子通道允许一些离子跨膜流过,而另一些离子(甚至 是那些与可跨膜流过离子密切相关的离子)不能通过,因此这些通道成为生物化学最迷人的 分子。这些门控离子通道在执行神经系统功能方面起中心作用。神经系统充当导线,允许精 细切换神经信号的快速流动。 最后,我们讨论另一种种通道,即细胞与细胞之间的通道(或缝隙连接,gap-junction)。 这种通道允许离子或代谢物质在细胞之间进行运输。例如分析连接的细胞间物质运输负责心 脏跳动时肌肉细胞同步收缩。 运输器的表达在很大程度上确定了特定细胞的代谢活性 各种细胞表达一套独有的细胞质膜运输器。由于这些运输器在很大程度上确定了细胞从 环境摄取离子和代谢物的模式,因此细胞表达的运输器组合非常重要。有些情况下,一个细 胞所表达的运输器组合就决定了这个细胞的性质,因为这个细胞只能摄取特定底物从而只能 执行特定的生物化学反应。 葡萄糖代谢能用来解释这种观点。如同我们在第 16 章葡萄糖代谢部分将详细讨论的, 组织之间的差异在于他们利用不同分子充当能源的能力。哪个组织能够利用葡萄糖主要取决 于它们表达葡萄糖运输器的情况。这些葡萄糖运输器包括 GLUT1, GLUT2, GLUT3, GLUT4, 和 GLUT5。例如 GLUT3 只在神经元和集中其它类型的细胞表达。在葡萄糖浓度相对较低 时,GLUT3 与葡萄糖结合相对较紧。在控制和整合活体代谢方面,这些运输器的表达非常
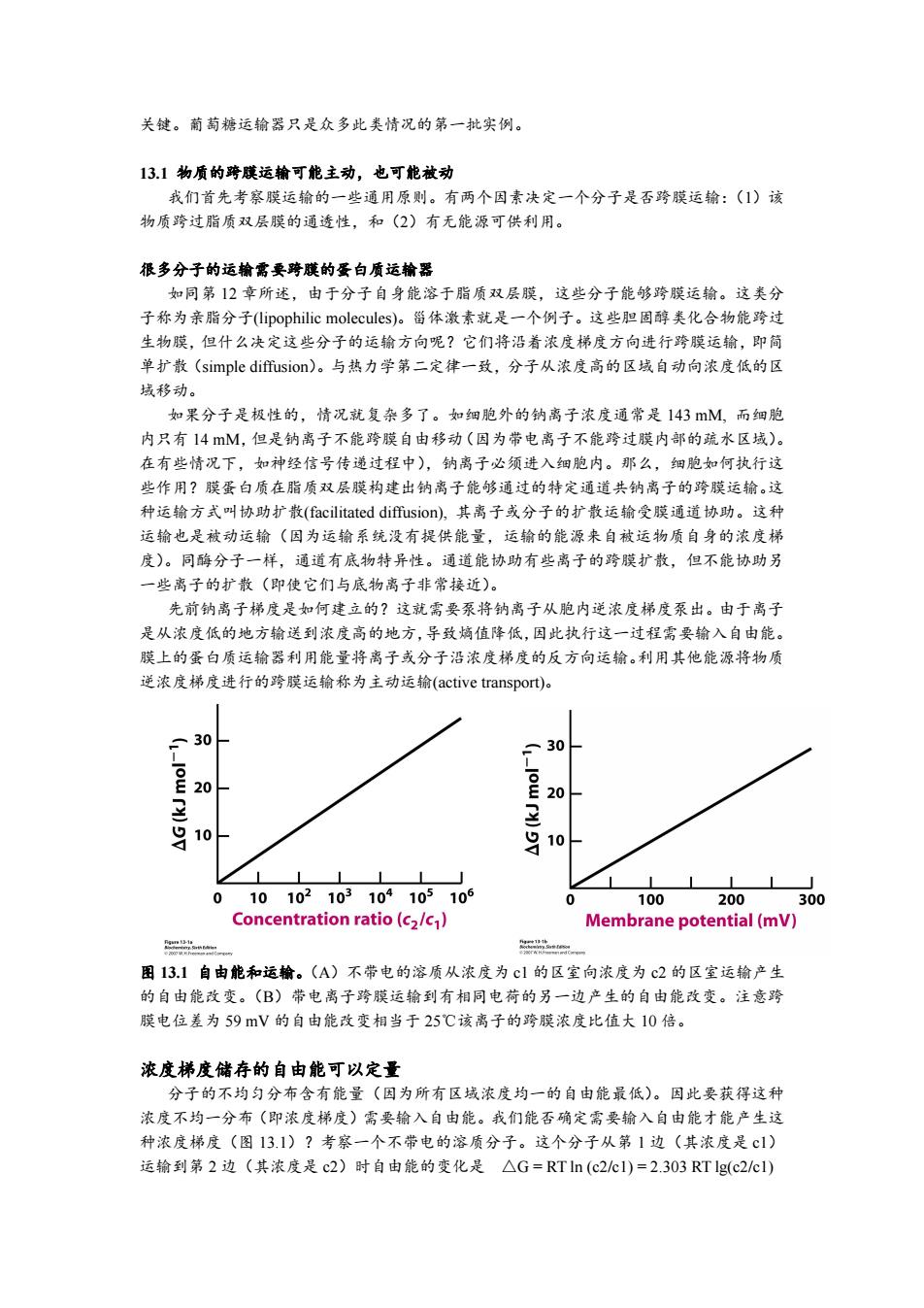
关键。葡萄糖运输器只是众多此类情况的第一批实例。 13.1物质的跨膜运输可能主动,也可能被动 我们首先考察膜运输的一些通用原则。有两个因素决定一个分子是否跨膜运输:(1)该 物质跨过脂质双层膜的通透性,和(2)有无能源可供利用。 很多分子的运输需要跨膜的蛋白质运输器 如同第12章所述,由于分子自身能溶于脂质双层膜,这些分子能够跨膜运输。这类分 子称为亲脂分子((lipophilic molecules)。甾体激素就是一个例子。这些胆固醇类化合物能跨过 生物膜,但什么决定这些分子的运输方向呢?它们将沿着浓度梯度方向进行跨膜运输,即简 单扩散(simple diffusion)。与热力学第二定律一致,分子从浓度高的区域自动向浓度低的区 域移动。 如果分子是极性的,情况就复杂多了。如细胞外的钠离子浓度通常是143mM,而细胞 内只有14M,但是钠离子不能跨膜自由移动(因为带电离子不能跨过膜内部的疏水区域)。 在有些情况下,如神经信号传递过程中),钠离子必须进入细胞内。那么,细胞如何执行这 些作用?膜蛋白质在脂质双层膜构建出钠离子能够通过的特定通道共钠离子的跨膜运输。这 种运输方式叫协助扩散(facilitated diffusion),其离子或分子的扩散运输受膜通道协助。这种 运输也是被动运输(因为运输系统没有提供能量,运输的能源来自被运物质自身的浓度梯 度)。同酶分子一样,通道有底物特异性。通道能协助有些离子的跨膜扩散,但不能协助另 一些离子的扩散(即使它们与底物离子非常接近)。 先前钠离子梯度是如何建立的?这就需要泵将钠离子从胞内逆浓度梯度泵出。由于离子 是从浓度低的地方输送到浓度高的地方,导致熵值降低,因此执行这一过程需要输入自由能。 膜上的蛋白质运输器利用能量将离子或分子沿浓度梯度的反方向运输。利用其他能源将物质 逆浓度梯度进行的跨膜运输称为主动运输(active transport)。 30 30 E20 显 10 吕10 E山 010102103104105106 100 200 300 Concentration ratio(c2/c) Membrane potential(mV) n 图13.1自由能和运输。(A)不带电的溶质从浓度为c1的区室向浓度为c2的区室运输产生 的自由能改变。(B)带电离子跨膜运输到有相同电荷的另一边产生的自由能改变。注意跨 膜电位差为59mV的自由能改变相当于25℃该离子的跨膜浓度比值大10倍。 浓度梯度储存的自由能可以定量 分子的不均匀分布含有能量(因为所有区域浓度均一的自由能最低)。因此要获得这种 浓度不均一分布(即浓度梯度)需要输入自由能。我们能否确定需要输入自由能才能产生这 种浓度梯度(图13.1)?考察一个不带电的溶质分子。这个分子从第1边(其浓度是c1) 运输到第2边(其浓度是c2)时自由能的变化是△G=RTln(c2cI)=2.303RTlg(c2/cl)
关键。葡萄糖运输器只是众多此类情况的第一批实例。 13.1 物质的跨膜运输可能主动,也可能被动 我们首先考察膜运输的一些通用原则。有两个因素决定一个分子是否跨膜运输:(1)该 物质跨过脂质双层膜的通透性,和(2)有无能源可供利用。 很多分子的运输需要跨膜的蛋白质运输器 如同第 12 章所述,由于分子自身能溶于脂质双层膜,这些分子能够跨膜运输。这类分 子称为亲脂分子(lipophilic molecules)。甾体激素就是一个例子。这些胆固醇类化合物能跨过 生物膜,但什么决定这些分子的运输方向呢?它们将沿着浓度梯度方向进行跨膜运输,即简 单扩散(simple diffusion)。与热力学第二定律一致,分子从浓度高的区域自动向浓度低的区 域移动。 如果分子是极性的,情况就复杂多了。如细胞外的钠离子浓度通常是 143 mM, 而细胞 内只有 14 mM,但是钠离子不能跨膜自由移动(因为带电离子不能跨过膜内部的疏水区域)。 在有些情况下,如神经信号传递过程中),钠离子必须进入细胞内。那么,细胞如何执行这 些作用?膜蛋白质在脂质双层膜构建出钠离子能够通过的特定通道共钠离子的跨膜运输。这 种运输方式叫协助扩散(facilitated diffusion), 其离子或分子的扩散运输受膜通道协助。这种 运输也是被动运输(因为运输系统没有提供能量,运输的能源来自被运物质自身的浓度梯 度)。同酶分子一样,通道有底物特异性。通道能协助有些离子的跨膜扩散,但不能协助另 一些离子的扩散(即使它们与底物离子非常接近)。 先前钠离子梯度是如何建立的?这就需要泵将钠离子从胞内逆浓度梯度泵出。由于离子 是从浓度低的地方输送到浓度高的地方,导致熵值降低,因此执行这一过程需要输入自由能。 膜上的蛋白质运输器利用能量将离子或分子沿浓度梯度的反方向运输。利用其他能源将物质 逆浓度梯度进行的跨膜运输称为主动运输(active transport)。 图 13.1 自由能和运输。(A)不带电的溶质从浓度为 c1 的区室向浓度为 c2 的区室运输产生 的自由能改变。(B)带电离子跨膜运输到有相同电荷的另一边产生的自由能改变。注意跨 膜电位差为 59 mV 的自由能改变相当于 25℃该离子的跨膜浓度比值大 10 倍。 浓度梯度储存的自由能可以定量 分子的不均匀分布含有能量(因为所有区域浓度均一的自由能最低)。因此要获得这种 浓度不均一分布(即浓度梯度)需要输入自由能。我们能否确定需要输入自由能才能产生这 种浓度梯度(图 13.1)?考察一个不带电的溶质分子。这个分子从第 1 边(其浓度是 c1) 运输到第 2 边(其浓度是 c2)时自由能的变化是 △G = RT ln (c2/c1) = 2.303 RT lg(c2/c1)

其中R是气体常数3.315x103kJ/mol,或1.987x103kcal/mol,T是绝对温度,单位是 kelvins。.对于带电离子跨膜不均匀分布还要考虑跨膜的电势差。浓度差和电势差的总和称为 膜电化学势能或膜势能。其自由能是 △G=RTln(c2cl)+ZF△V=2.303RTlg(c2/cl)+ZF△V 其中Z是被运离子的电荷,△V是跨膜电势差,F是法拉利常数(96.5kJN.mol,或23.1kcal V.mol)。 如果△G是正值,运输是主动的。如果△G是负值,运输时被动的。例如,不带电物质 从cl=103M运输到c2=10-M,△G=2.303RTlg(10-10)=+11.4 kJ/mol(若温度是 298k)。这个数值表明,这种运输需要能量,是主动运输。 13.2有两类膜蛋白利用ATP水解将离子或分子泵过膜 动物细胞的胞外液体的盐浓度与海水相似。但是细胞必须控制细胞内离子浓度,防止离 子达到有害的高浓度状态,有利于一些生化反应的进行。例如,与胞外溶液相比,大多数动 物细胞内含有高浓度的钾离子,但钠离子浓度较低。这些离子浓度梯度是特定的运输系统造 成的。这个运输系统是Na*-K+泵或Na*-K+ATPase。.这个泵对ATP的水解产生了Na*主 动运出细胞和K+主动运入细胞所需要的能量,产生离子梯度。只有N和K+都存在时, 这个泵才能水解ATP,所以称为Na-K+ATPase。与其它ATPase一样,该酶需要Mg+. Na和K+运输伴随着自由能的变化可以计算。假定胞外钠离子浓度是143mM,胞内是 14mM,而胞外钾离子浓度是4mM,胞内是157mM,跨膜电位是-50mV,温度是37℃, 那么运输3 mole Na+到胞外,运输2 mole K+到胞内需要3x5.99+2x9.46=+36.9kJ/mol。 ATP水解能提供50kJ/mol来驱动离子的主动运输。Na*和K+主动运输有重大的生理意义。 实际上没有运动的细胞内超过1/3的ATP用来驱动这些离子泵的运转。动物细胞的Na和 K浓度梯度控制细胞体积,使神经元和肌肉细胞能被电激活,驱动糖和氨基酸的主动运输。 随后纯化的其它离子泵表明离子泵蛋白质存在于从细菌、古生菌、到所有真核生物,是 一个蛋白大家族。这些离子泵能够主动运输特定离子。其中两个泵有特殊意义。Ca+ATPase 能够将Ca+运出细胞质,运入肌细胞内质网:胃H-K+ATPase将质子泵入胃,使之pH值 低于l.0。这些酶和众多已知的同源物,包括Na-K+ATPase,称为P-类ATPase,因为它们 能形成关键的磷酸化中间产物。在形成该中间产物的过程中,ATP提供的磷酸基团与ATPase 的一个保守的天冬氨酸残基的侧链形成共价连接。 Phosphorylaspartate
其中 R 是气体常数 3.315 x 10 -3 kJ/mol, 或 1.987 x 10 -3 kcal / mol, T 是绝对温度,单位是 kelvins。对于带电离子跨膜不均匀分布还要考虑跨膜的电势差。浓度差和电势差的总和称为 膜电化学势能或膜势能。其自由能是 △G = RT ln (c2/c1) + ZF △V = 2.303 RT lg(c2/c1) + ZF △V 其中 Z 是被运离子的电荷,△V 是跨膜电势差,F 是法拉利常数(96.5 kJ/V.mol, 或 23.1 kcal /V.mol)。 如果△G 是正值,运输是主动的。如果△G 是负值,运输时被动的。例如,不带电物质 从 c1 = 10 -3M 运输到 c2 = 10 -1M, △G = 2.303 RT lg (10 -1 /10 -3) = + 11.4 kJ/mol (若温度是 298k)。这个数值表明,这种运输需要能量,是主动运输。 13.2 有两类膜蛋白利用 ATP 水解将离子或分子泵过膜 动物细胞的胞外液体的盐浓度与海水相似。但是细胞必须控制细胞内离子浓度,防止离 子达到有害的高浓度状态,有利于一些生化反应的进行。例如,与胞外溶液相比,大多数动 物细胞内含有高浓度的钾离子,但钠离子浓度较低。这些离子浓度梯度是特定的运输系统造 成的。这个运输系统是 Na + - K+ 泵或 Na + - K+ ATPase。这个泵对 ATP 的水解产生了 Na + 主 动运出细胞和 K+主动运入细胞所需要的能量,产生离子梯度。只有 Na + 和 K+都存在时, 这个泵才能水解 ATP,所以称为 Na + - K+ ATPase。与其它 ATPase 一样,该酶需要 Mg 2+ . Na +和 K+运输伴随着自由能的变化可以计算。假定胞外钠离子浓度是 143 mM,胞内是 14 mM,而胞外钾离子浓度是 4 mM, 胞内是 157 mM, 跨膜电位是-50 mV,温度是 37℃, 那么运输 3 mole Na+到胞外,运输 2 mole K+到胞内需要 3 x 5.99 + 2 x 9.46 = + 36.9 kJ/mol。 ATP 水解能提供 50 kJ/mol 来驱动离子的主动运输。Na +和 K+主动运输有重大的生理意义。 实际上没有运动的细胞内超过 1/3 的 ATP 用来驱动这些离子泵的运转。动物细胞的 Na +和 K+浓度梯度控制细胞体积,使神经元和肌肉细胞能被电激活,驱动糖和氨基酸的主动运输。 随后纯化的其它离子泵表明离子泵蛋白质存在于从细菌、古生菌、到所有真核生物,是 一个蛋白大家族。这些离子泵能够主动运输特定离子。其中两个泵有特殊意义。Ca 2+ ATPase 能够将 Ca 2+运出细胞质,运入肌细胞内质网;胃 H+ -K+ ATPase 将质子泵入胃,使之 pH 值 低于 1.0。这些酶和众多已知的同源物,包括 Na + - K+ ATPase,称为 P-类 ATPase,因为它们 能形成关键的磷酸化中间产物。在形成该中间产物的过程中,ATP 提供的磷酸基团与 ATPase 的一个保守的天冬氨酸残基的侧链形成共价连接

P-类ATPase将磷酸化和泵构象转换偶联,实现钙离子的跨膜运输 膜泵作用原理简单,但是作用细节很复杂。基本上,每个膜泵蛋白质有两种功能状态, 即朝向生物膜一侧开放的离子结合状态和朝向生物膜另一侧开放的离子结合状态(图13.2)。 为了在单一方向跨膜运输离子,自由能的利用必须与泵蛋白构象转换偶联。 Energy input Conformation 1 Conformation 2 图132泵作用。用膜泵将一个分子跨膜运输的示意图。泵在两种构型之间相互转换,每种 构型都有一个离子结合位点,但是泵的开口分别处于生物膜的不同侧面。 我们以肌肉细胞内质网膜的Ca2+ATPase为例,来介绍P-类ATPase结构域特征。依靠5 种不同状态泵的晶体结构,已经确定了这类ATPase成员的详细特性。肌肉细胞内质网膜的 Ca2+ATPase站内质网膜蛋白量的80%在肌肉收缩方面起重要作用。细胞质Ca+浓度迅速提 升导致肌肉收缩。而细胞质C+迅速运入内质网导致细胞质Ca+浓度迅速降低使肌肉松弛。 度迅速提升导致肌肉收缩。肌细胞内质网是储存Ca+的特定场所。将细胞质Ca+运入肌细 胞内质网的是SERCA。这种泵使细胞质Ca+浓度维持在0.1M,而内质网的Ca+浓度处 于1.5mM水平。 阐明的第一例SERCA有Ca+结合,但没有核苷酸(图13.3)。SERCA只有一条多肽链, 大小是110kD,有一个含有10个螺旋的跨膜区。这个跨膜区有两个C+结合位点。每个 C+能够与7个氧原子形成配位键。这7个氧原子来自多肽链的谷氨酸、天冬氨酸、苏氨酸、 天冬酰胺侧链基团,多肽链骨架的羰基,和水分子。面向细胞质的头部大,相当于整个蛋白 质质量的一半,由三个不同的结构域构成。这三个结构域有不同的功能。一个结构域(称为 N)与ATP结合,另一个结构域(称为P)的保守天冬氨酸接受磷酸,第三个结构域(A)充 当调解器(actuator),将蛋白质N和P结构域的变化与这个蛋白的跨膜区域联系起来。 bb304 Asn 798 bb307 Glu 308 图13.3Ca2+泵结构。P.类ATPase成员SERCA的结构。注意两个钙离子(绿色)处于跨膜 结构域的中心。保守的天冬氨酸残基(As即351)与磷酸基团结合,位于P结构域。bb表示多 肽骨架链的羰基
P-类 ATPase 将磷酸化和泵构象转换偶联,实现钙离子的跨膜运输 膜泵作用原理简单,但是作用细节很复杂。基本上,每个膜泵蛋白质有两种功能状态, 即朝向生物膜一侧开放的离子结合状态和朝向生物膜另一侧开放的离子结合状态(图 13.2)。 为了在单一方向跨膜运输离子,自由能的利用必须与泵蛋白构象转换偶联。 图 13.2 泵作用。用膜泵将一个分子跨膜运输的示意图。泵在两种构型之间相互转换,每种 构型都有一个离子结合位点,但是泵的开口分别处于生物膜的不同侧面。 我们以肌肉细胞内质网膜的 Ca 2+ ATPase 为例,来介绍 P-类 ATPase 结构域特征。依靠 5 种不同状态泵的晶体结构,已经确定了这类 ATPase 成员的详细特性。肌肉细胞内质网膜的 Ca 2+ ATPase 站内质网膜蛋白量的 80%在肌肉收缩方面起重要作用。细胞质 Ca 2+浓度迅速提 升导致肌肉收缩。而细胞质 Ca 2+迅速运入内质网导致细胞质 Ca 2+浓度迅速降低使肌肉松弛。 度迅速提升导致肌肉收缩。肌细胞内质网是储存 Ca 2+的特定场所。将细胞质 Ca 2+运入肌细 胞内质网的是 SERCA。这种泵使细胞质 Ca 2+浓度维持在 0.1 M,而内质网的 Ca 2+浓度处 于 1.5 mM 水平。 阐明的第一例 SERCA 有 Ca 2+结合,但没有核苷酸(图 13.3)。SERCA 只有一条多肽链, 大小是 110 kD,有一个含有 10 个 螺旋的跨膜区。这个跨膜区有两个 Ca 2+结合位点。每个 Ca 2+能够与 7 个氧原子形成配位键。这 7 个氧原子来自多肽链的谷氨酸、天冬氨酸、苏氨酸、 天冬酰胺侧链基团,多肽链骨架的羰基,和水分子。面向细胞质的头部大,相当于整个蛋白 质质量的一半,由三个不同的结构域构成。这三个结构域有不同的功能。一个结构域(称为 N)与 ATP 结合,另一个结构域(称为 P)的保守天冬氨酸接受磷酸,第三个结构域(A)充 当调解器(actuator),将蛋白质 N 和 P 结构域的变化与这个蛋白的跨膜区域联系起来。 图 13.3 Ca 2+泵结构。P-类 ATPase 成员 SERCA 的结构。注意两个钙离子(绿色)处于跨膜 结构域的中心。保守的天冬氨酸残基(Asp351)与磷酸基团结合,位于 P 结构域。bb 表示多 肽骨架链的羰基
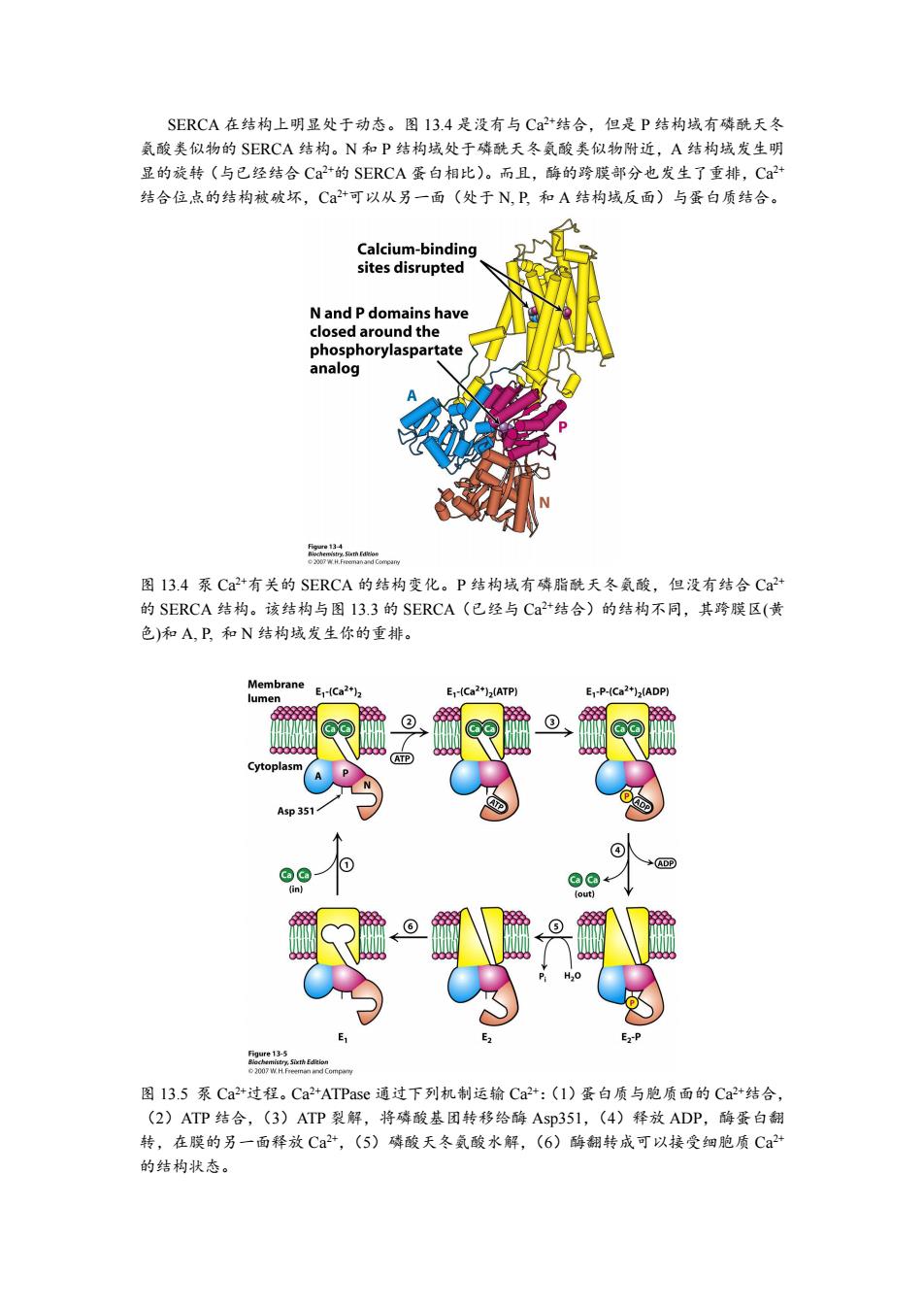
SERCA在结构上明显处于动态。图13.4是没有与C2+结合,但是P结构域有磷酰天冬 氨酸类似物的SERCA结构。N和P结构域处于磷酰天冬氨酸类似物附近,A结构域发生明 显的旋转(与已经结合Ca+的SERCA蛋白相比)。而且,酶的跨膜部分也发生了重排,Ca+ 结合位点的结构被破坏,C+可以从另一面(处于N,P,和A结构域反面)与蛋白质结合。 Calcium-binding sites disrupted N and P domains have closed around the phosphorylaspartate analog 2sfsgCapaw 图13.4泵Ca2+有关的SERCA的结构变化。P结构域有磷脂酰天冬氨酸,但没有结合Ca2+ 的SERCA结构。该结构与图13.3的SERCA(已经与Ca+结合)的结构不同,其跨膜区(黄 色)和A,P,和N结构域发生你的重排。 Membrane E1-Ca2+)2 lumen E-(Ca2+)(ATP) E-P-(Ca2+)2(ADP) ② ATP) Cytoplasm ⊙ 4sp35 ④ 1 ADP @©← (in) fout] E1 E2 E-P Figure 13-5 p 图13.5泵Ca2+过程。Ca2+ATPase通过下列机制运输Ca2+:(1)蛋白质与胞质面的Ca2+结合, (2)ATP结合,(3)ATP裂解,将磷酸基团转移给酶Asp351,(4)释放ADP,酶蛋白翻 转,在膜的另一面释放Ca2+,(5)磷酸天冬氨酸水解,(6)酶翻转成可以接受细胞质Ca2+ 的结构状态
SERCA 在结构上明显处于动态。图 13.4 是没有与 Ca 2+结合,但是 P 结构域有磷酰天冬 氨酸类似物的 SERCA 结构。N 和 P 结构域处于磷酰天冬氨酸类似物附近,A 结构域发生明 显的旋转(与已经结合 Ca 2+的 SERCA 蛋白相比)。而且,酶的跨膜部分也发生了重排,Ca 2+ 结合位点的结构被破坏,Ca 2+可以从另一面(处于 N, P, 和 A 结构域反面)与蛋白质结合。 图 13.4 泵 Ca 2+有关的 SERCA 的结构变化。P 结构域有磷脂酰天冬氨酸,但没有结合 Ca 2+ 的 SERCA 结构。该结构与图 13.3 的 SERCA(已经与 Ca 2+结合)的结构不同,其跨膜区(黄 色)和 A, P, 和 N 结构域发生你的重排。 图 13.5 泵 Ca 2+过程。Ca 2+ATPase 通过下列机制运输 Ca 2+:(1)蛋白质与胞质面的 Ca 2+结合, (2)ATP 结合,(3)ATP 裂解,将磷酸基团转移给酶 Asp351,(4)释放 ADP,酶蛋白翻 转,在膜的另一面释放 Ca 2+,(5)磷酸天冬氨酸水解,(6)酶翻转成可以接受细胞质 Ca 2+ 的结构状态