
第五章基因和基因组研究技术 从caterpillar发展成蝴蝶,基因表达谱发生了一系列显著变化。用DNA芯片能同时检测成 千上万个基因的表达水平。右边是一张基因芯片,显示人的12000多种基因表达水平。每个 点的量度表示该基因的表达强度。 自从1970年代出现DNA重组技术以来,重组DNA技术给生物化学带来了革命性进步。 生物个体的遗传特征能够根据人类设计加以改造。重组DNA技术是人类对DNA,RNA, 和病毒研究进行了几十年研究的结晶。该技术取决于酶切、连接、和复制DNA的酶和RNA 反转录。限制性内切酶能够将很长的DNA分子特异断裂成易于操作的DNA片段:DNA连 接酶能够将DNA片段连接。现在已经有很多限制性内切酶。采用这种技术,研究人员能够 将将DNA片段从一个DNA分子转移到另一DNA分子中。因此重组DNA技术的基础是核 酸酶学 第二个基础是碱基配对,即序列互补的核酸链相互间自动识别、自动结合形成核酸双链。 用互补DNA或RNA探针杂交检测特异核酸序列灵敏度高。在重组DNA技术中,碱基配对 用于重组DNA构建、检测和扩增特异核酸序列。重组DNA技术也依赖病毒。病毒能有效 地将自身DNA(或RNA)供应宿主,挟持宿主复制病毒基因组、合成病毒蛋白质,或将病毒 DNA整合到宿主基因组中。质粒是细菌染色体外的遗传物质,这种遗传物质对重组DNA 技术而言也是不可或缺的。 第三,已经建立了非常有效的DNA序列测定技术。这些技术能够测定全基因组序列。 最先测定的是病毒基因组,随后是更长的细菌基因组,最后测定的是真核基因组,包括30 亿碱基对长的人类基因组。科研人员正在开始探讨这些基因组序列的海量信息。 这些新技术极大地促进了相关领域的科学研究。即可以在天然环境中、也可以在其它条 件下研究某个基因或基因产物的作用。以某种方式改造基因,制造变异蛋白质能够详细了解 蛋白质的功能。重组DNA技术能够制造临床上有用的蛋白质,如激素:也能用来制造耐受 病虫害或恶劣生长环境的农作物。重组DNA技术提供的这些机会将产生更大的影响
第五章 基因和基因组研究技术 从 caterpillar 发展成蝴蝶,基因表达谱发生了一系列显著变化。用 DNA 芯片能同时检测成 千上万个基因的表达水平。右边是一张基因芯片,显示人的 12000 多种基因表达水平。每个 点的量度表示该基因的表达强度。 自从 1970 年代出现 DNA 重组技术以来,重组 DNA 技术给生物化学带来了革命性进步。 生物个体的遗传特征能够根据人类设计加以改造。重组 DNA 技术是人类对 DNA,RNA, 和病毒研究进行了几十年研究的结晶。该技术取决于酶切、连接、和复制 DNA 的酶和 RNA 反转录。限制性内切酶能够将很长的 DNA 分子特异断裂成易于操作的 DNA 片段;DNA 连 接酶能够将 DNA 片段连接。现在已经有很多限制性内切酶。采用这种技术,研究人员能够 将将 DNA 片段从一个 DNA 分子转移到另一 DNA 分子中。因此重组 DNA 技术的基础是核 酸酶学 第二个基础是碱基配对,即序列互补的核酸链相互间自动识别、自动结合形成核酸双链。 用互补 DNA 或 RNA 探针杂交检测特异核酸序列灵敏度高。在重组 DNA 技术中,碱基配对 用于重组 DNA 构建、检测和扩增特异核酸序列。重组 DNA 技术也依赖病毒。病毒能有效 地将自身 DNA(或 RNA)供应宿主,挟持宿主复制病毒基因组、合成病毒蛋白质,或将病毒 DNA 整合到宿主基因组中。质粒是细菌染色体外的遗传物质,这种遗传物质对重组 DNA 技术而言也是不可或缺的。 第三,已经建立了非常有效的 DNA 序列测定技术。这些技术能够测定全基因组序列。 最先测定的是病毒基因组,随后是更长的细菌基因组,最后测定的是真核基因组,包括 30 亿碱基对长的人类基因组。科研人员正在开始探讨这些基因组序列的海量信息。 这些新技术极大地促进了相关领域的科学研究。即可以在天然环境中、也可以在其它条 件下研究某个基因或基因产物的作用。以某种方式改造基因,制造变异蛋白质能够详细了解 蛋白质的功能。重组 DNA 技术能够制造临床上有用的蛋白质,如激素;也能用来制造耐受 病虫害或恶劣生长环境的农作物。重组 DNA 技术提供的这些机会将产生更大的影响

5.1基因研究的关键工具 生物技术的迅猛发展实际上是几个关键技术的产物 1.限制性内切酶分析。限制性内切酶是精确的剪切DNA分子的手术刀,使研究人员能够 操纵DNA片段。 2.印迹技术。Southern blots和Northern blots能分别用来分离和鉴定DNA和RNA。在前 面介绍的Western blots采用抗体鉴定蛋白质。 3.DNA序列测定。该技术能够测定DNA分子的碱基序列。DNA序列测定产生了大量的 信息,这些信息涉及基因结构、基因表达调控、和蛋白质结构。 4.核酸固相合成。能够化学合成所需序列的核酸分子用于鉴定或扩增其它核酸。 5.聚合酶链式反应(PCR)。PCR能使特定DNA片段扩增上十亿倍。PCR反应能够将一个 DNA分子扩增到足以进行DNA鉴定和操作的产量。该技术用于病原体鉴定和遗传病 诊断、确定犯罪现场毛发来源、复活化石的基因。 6.最后的一套技术依赖计算机,将在第6章介绍。没有计算机技术,我们不可能对大量信 息(尤其是DNA序列信息)进行分类、检索、和鉴定。而这些信息就是用前面提到的 那些技术获得的。 限制性内切酶能够将DNA分子断裂成特异片段 限制酶也叫限制性核酸内切酶,能够识别双链DNA的特定碱基序列并在特定位点断裂 DNA双链。这种精确的分子手术刀是大自然馈赠给生物化学工作者的礼物。分析染色体结 构、测定非常长的DNA序列、分离基因、制造能被克隆的DNA分子都离不开限制性内切 酶。在l960年代后期,Werner Alber和Hamilton Smith发现了限制性内切酶,Daniel Natherns 首先应用限制性内切酶。 不同原核生物都有限制性内切酶。限制性内切酶的生物功能是断裂外源DNA分子,但 由于限制酶识别位点的碱基序列已被甲基化,因此限制酶不断裂生物自身的DNA分子。很 多限制酶识别的核酸序列长度是4~8个碱基对,这个区域两条核酸链都会被这个核酸内切 酶水解。这些酶识别序列的显著特征是多数限制酶识别位点具有二重旋转对称。换句话说, 使回文结构,或倒置重复。限制性内切酶断裂位点也处于核酸链的对称位置。例如,来源于 Streptomyces achromogenes的限制性内切酶识别的核苷酸序列是 Cleavage site ↓ 5'C一C一G一C一G一G3' 3' G-G-C+G-c-c 5 Cleavage site Symmetry axis 这个酶水解对称轴两侧的C-G磷酸二酯键。在第9章所介绍的内容指出,限制性核酸 内切酶酶切位,点的对称性是由核酸内切酶自身结构决定的。 现在已经鉴定并纯化的限制性内切酶有几百种。对这些限制性内切酶的命名方案是前面 的三个斜写字母表示限制性内切酶的菌种来源(例如Eco表示限制酶来自Esterichia coli,Hin 表示限制酶来自Haemophilus influenzae,Hae表示限制酶来自Haemophilus aegyptius),后面 是菌株名(如有必要)和罗马数字(如果同一菌株分离出一种以上的限制性内切酶)。图51 列出了几个限制性内切酶的酶切位点
5.1 基因研究的关键工具 生物技术的迅猛发展实际上是几个关键技术的产物 1. 限制性内切酶分析。限制性内切酶是精确的剪切 DNA 分子的手术刀,使研究人员能够 操纵 DNA 片段。 2. 印迹技术。Southern blots 和 Northern blots 能分别用来分离和鉴定 DNA 和 RNA。在前 面介绍的 Western blots 采用抗体鉴定蛋白质。 3. DNA 序列测定。该技术能够测定 DNA 分子的碱基序列。DNA 序列测定产生了大量的 信息,这些信息涉及基因结构、基因表达调控、和蛋白质结构。 4. 核酸固相合成。能够化学合成所需序列的核酸分子用于鉴定或扩增其它核酸。 5. 聚合酶链式反应(PCR)。PCR 能使特定 DNA 片段扩增上十亿倍。PCR 反应能够将一个 DNA 分子扩增到足以进行 DNA 鉴定和操作的产量。该技术用于病原体鉴定和遗传病 诊断、确定犯罪现场毛发来源、复活化石的基因。 6. 最后的一套技术依赖计算机,将在第 6 章介绍。没有计算机技术,我们不可能对大量信 息(尤其是 DNA 序列信息)进行分类、检索、和鉴定。而这些信息就是用前面提到的 那些技术获得的。 限制性内切酶能够将 DNA 分子断裂成特异片段 限制酶也叫限制性核酸内切酶,能够识别双链 DNA 的特定碱基序列并在特定位点断裂 DNA 双链。这种精确的分子手术刀是大自然馈赠给生物化学工作者的礼物。分析染色体结 构、测定非常长的 DNA 序列、分离基因、制造能被克隆的 DNA 分子都离不开限制性内切 酶。在 1960 年代后期,Werner Alber 和 Hamilton Smith 发现了限制性内切酶,Daniel Natherns 首先应用限制性内切酶。 不同原核生物都有限制性内切酶。限制性内切酶的生物功能是断裂外源 DNA 分子,但 由于限制酶识别位点的碱基序列已被甲基化,因此限制酶不断裂生物自身的 DNA 分子。很 多限制酶识别的核酸序列长度是 4 ~ 8 个碱基对,这个区域两条核酸链都会被这个核酸内切 酶水解。这些酶识别序列的显著特征是多数限制酶识别位点具有二重旋转对称。换句话说, 使回文结构,或倒置重复。限制性内切酶断裂位点也处于核酸链的对称位置。例如,来源于 Streptomyces achromogenes 的限制性内切酶识别的核苷酸序列是 这个酶水解对称轴两侧的 C-G 磷酸二酯键。在第 9 章所介绍的内容指出,限制性核酸 内切酶酶切位点的对称性是由核酸内切酶自身结构决定的。 现在已经鉴定并纯化的限制性内切酶有几百种。对这些限制性内切酶的命名方案是前面 的三个斜写字母表示限制性内切酶的菌种来源(例如 Eco 表示限制酶来自 Esterichia coli, Hin 表示限制酶来自 Haemophilus influenzae,Hae 表示限制酶来自 Haemophilus aegyptius),后面 是菌株名(如有必要)和罗马数字(如果同一菌株分离出一种以上的限制性内切酶)。图 5.1 列出了几个限制性内切酶的酶切位点

5'GGATCC 3' BamHI 3'CCTAGG 5' 个 5'GAATTC 3' EcoRI 3'CTTAAG 5' ↓ 5'GGCC 3 Haelll 3'CCGG 5 ↑ ↓ 5'GCGC 3 Hhal 3'CGCG 5 5'CTCGAG3' Xhol 3'GAGCTC5' Figures. ry.Sixth Edition 2007 W.H.Freeman and Compary 图51一些限制性内切酶的特异性。这些酶识别序列有二重对称轴。围绕该对称轴(绿色) 旋转180度,序列与原始序列相同。红色箭头标出限制酶的酶切位置。右边是限制酶的缩写 名称。注意:有些限制酶平切,有些限制酶错开切(产生粘性末端)。 限制性内切酶能够将DNA分子断裂成易于进行分析和操纵的特异片段。例如长度为 5.1kb的SV40是双链环状DNA,能产生肿瘤。这个DNA分子有一个EcoRI切点,4个Hpal 切点,和I1个HimdⅢ切点。一个限制性内切酶酶切DNA的产物可以用另一个限制性内切 酶酶切产生更小的DNA片段。DNA分子为限制性内切酶酶切产生的DNA片段图谱可以作 为该DNA分子的指纹。实际上,用一些列限制型内切酶酶切产生的限制型内切酶图谱可以 对复杂染色体进行作图(这样的染色体可能含有数亿碱基对)。 凝胶电泳分离、检测限制酶酶切的DNA片段 由于凝胶电泳能够分离、显示限制酶酶切的DNA片段,因此能够检测DNA分子间的 细小差异。凝胶电泳的种类很多,大多数凝胶电泳的迁移率与DNA分子式量的对数值呈负 线性关系(在一定的分子式量范围内)。聚丙烯酰胺凝胶的范围达到1000碱基对,而琼脂糖 凝胶孔径更大,分子范围达到20b。这些凝胶电泳的一个典型特征是分辨率高有些凝胶电 泳条件能够区分在几百个碱基对长的DNA分子间只相差一个核苷酸的DNA片段。而且长 度达到数百万碱基对长度的整条染色体也能用琼脂糖凝胶电泳分离。此时采用的电场是在不 同方向施加脉冲电场(即脉冲电场凝胶电泳,P℉GE)。染色体在拉伸和松弛方面有差异,因 此短期内施加和切断电场能够在琼脂糖凝胶上分离染色体。放射性自显影能够显示凝胶上同 位素标记的DNA样品条带或点。另外,用溴化乙锭染色也能显示凝胶上的DNA分子。当 溴化乙锭插入DNA双螺旋,DNA分子显示强烈的橘红色荧光(图5.2)。一个条带只有50ng 的DNA量就足以用溴化乙锭显色观察
图 5.1 一些限制性内切酶的特异性。这些酶识别序列有二重对称轴。围绕该对称轴(绿色) 旋转 180 度,序列与原始序列相同。红色箭头标出限制酶的酶切位置。右边是限制酶的缩写 名称。注意:有些限制酶平切,有些限制酶错开切(产生粘性末端)。 限制性内切酶能够将 DNA 分子断裂成易于进行分析和操纵的特异片段。例如长度为 5.1kb 的 SV40 是双链环状 DNA,能产生肿瘤。这个 DNA 分子有一个 EcoRI 切点,4 个 HpaI 切点,和 11 个 HindIII 切点。一个限制性内切酶酶切 DNA 的产物可以用另一个限制性内切 酶酶切产生更小的 DNA 片段。DNA 分子为限制性内切酶酶切产生的 DNA 片段图谱可以作 为该 DNA 分子的指纹。实际上,用一些列限制型内切酶酶切产生的限制型内切酶图谱可以 对复杂染色体进行作图(这样的染色体可能含有数亿碱基对)。 凝胶电泳分离、检测限制酶酶切的 DNA 片段 由于凝胶电泳能够分离、显示限制酶酶切的 DNA 片段,因此能够检测 DNA 分子间的 细小差异。凝胶电泳的种类很多,大多数凝胶电泳的迁移率与 DNA 分子式量的对数值呈负 线性关系(在一定的分子式量范围内)。聚丙烯酰胺凝胶的范围达到 1000 碱基对,而琼脂糖 凝胶孔径更大,分子范围达到 20kb。这些凝胶电泳的一个典型特征是分辨率高有些凝胶电 泳条件能够区分在几百个碱基对长的 DNA 分子间只相差一个核苷酸的 DNA 片段。而且长 度达到数百万碱基对长度的整条染色体也能用琼脂糖凝胶电泳分离。此时采用的电场是在不 同方向施加脉冲电场(即脉冲电场凝胶电泳,PFGE)。染色体在拉伸和松弛方面有差异,因 此短期内施加和切断电场能够在琼脂糖凝胶上分离染色体。放射性自显影能够显示凝胶上同 位素标记的 DNA 样品条带或点。另外,用溴化乙锭染色也能显示凝胶上的 DNA 分子。当 溴化乙锭插入 DNA 双螺旋,DNA 分子显示强烈的橘红色荧光(图 5.2)。一个条带只有 50 ng 的 DNA 量就足以用溴化乙锭显色观察
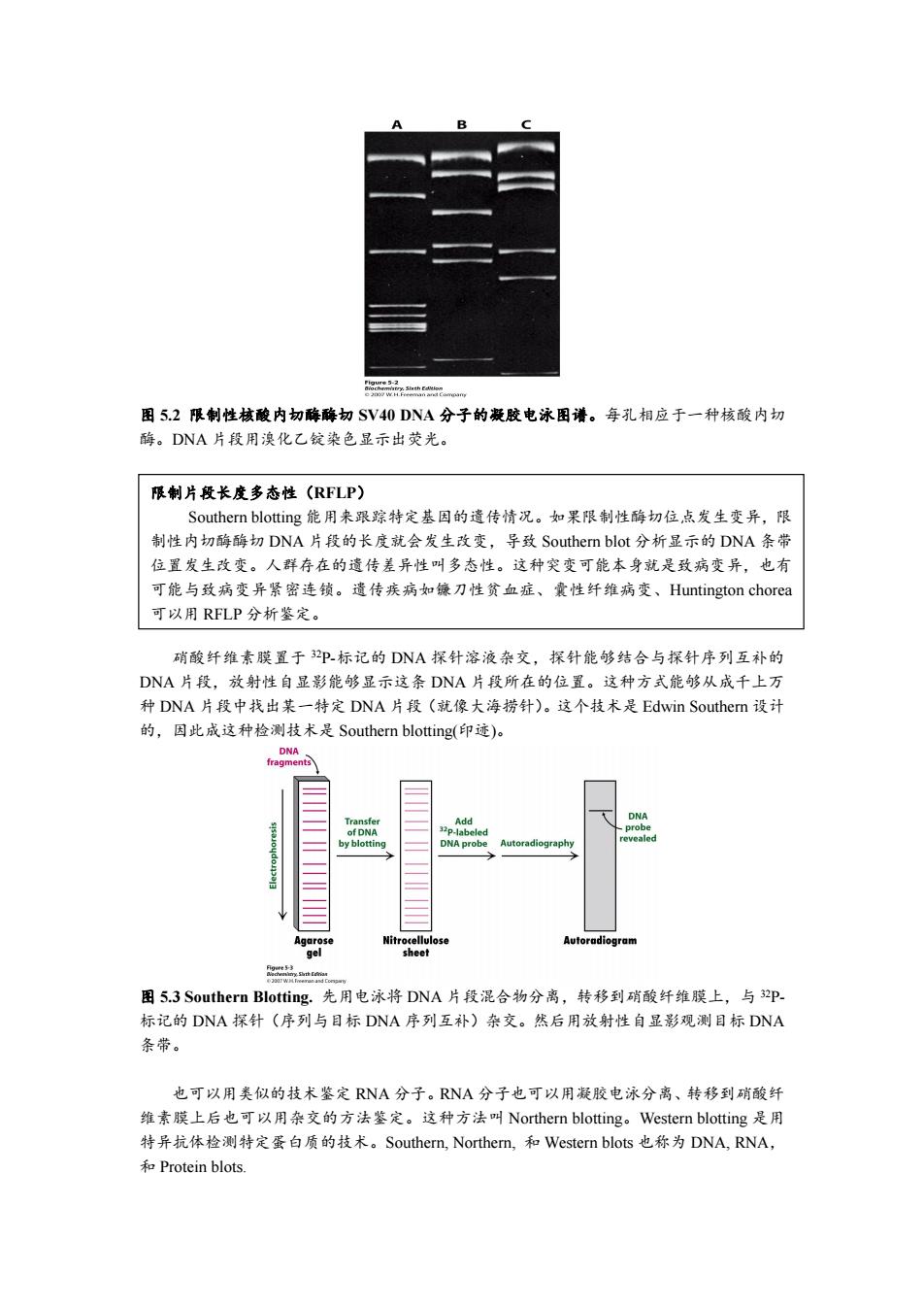
二品2n 图52限制性核酸内切酶酶切SV40DNA分子的凝胶电泳图谱。每孔相应于一种核酸内切 酶。DNA片段用溴化乙锭染色显示出荧光。 限制片段长度多态性(RFLP) Southern blotting能用来跟踪特定基因的遗传情况。如果限制性酶切位,点发生变异,限 制性内切酶酶切DNA片段的长度就会发生改变,导致Southern blot分析显示的DNA条带 位置发生改变。人群存在的遗传差异性叫多态性。这种突变可能本身就是致病变异,也有 可能与致病变异紧密连锁。遗传疾病如镰刀性贫血症、囊性纤维病变、Huntington chorea 可以用RFLP分析鉴定。 硝酸纤维素膜置于3P标记的DNA探针溶液杂交,探针能够结合与探针序列互补的 DNA片段,放射性自显影能够显示这条DNA片段所在的位置。这种方式能够从成千上万 种DNA片段中找出某一特定DNA片段(就像大海捞针)。这个技术是Edwin Southern设计 的,因此成这种检测技术是Southern blotting(印迹)。 DNA fragments e Add DNA -labeled DNA probe Autoradiography Agarose Nitrocellulo Autoradiogram gel sheet 品 图5.3 Southern Blotting.先用电泳将DNA片段混合物分离,转移到硝酸纤维膜上,与p 标记的DNA探针(序列与目标DNA序列互补)杂交。然后用放射性自显影观测目标DNA 条带。 也可以用类似的技术鉴定RNA分子。RNA分子也可以用凝胶电泳分离、转移到硝酸纤 维素膜上后也可以用杂交的方法鉴定。这种方法叫Northern blotting。Western blotting是用 特异抗体检测特定蛋白质的技术。Southern,Northern,和Western blots也称为DNA,RNA, 和Protein blots
图 5.2 限制性核酸内切酶酶切 SV40 DNA 分子的凝胶电泳图谱。每孔相应于一种核酸内切 酶。DNA 片段用溴化乙锭染色显示出荧光。 用标记的核酸探针杂交凝胶分离的限制型内切酶酶切条带,能够鉴定序列与探针互补的 DNA 片段(图 5.3)。限制性酶切产物用琼脂糖凝胶分离后,变性转化成单链 DNA,转移到 硝酸纤维素膜上。DNA 片段在硝酸纤维素膜的位置与原来琼脂糖凝胶的位置相同。然后将 硝酸纤维素膜置于 32P-标记的 DNA 探针溶液杂交,探针能够结合与探针序列互补的 DNA 片段,放射性自显影能够显示这条 DNA 片段所在的位置。这种方式能够从成千上万 种 DNA 片段中找出某一特定 DNA 片段(就像大海捞针)。这个技术是 Edwin Southern 设计 的,因此成这种检测技术是 Southern blotting(印迹)。 图 5.3 Southern Blotting. 先用电泳将 DNA 片段混合物分离,转移到硝酸纤维膜上,与 32P- 标记的 DNA 探针(序列与目标 DNA 序列互补)杂交。然后用放射性自显影观测目标 DNA 条带。 也可以用类似的技术鉴定 RNA 分子。RNA 分子也可以用凝胶电泳分离、转移到硝酸纤 维素膜上后也可以用杂交的方法鉴定。这种方法叫 Northern blotting。Western blotting 是用 特异抗体检测特定蛋白质的技术。Southern, Northern, 和 Western blots 也称为 DNA, RNA, 和 Protein blots. 限制片段长度多态性(RFLP) Southern blotting 能用来跟踪特定基因的遗传情况。如果限制性酶切位点发生变异,限 制性内切酶酶切 DNA 片段的长度就会发生改变,导致 Southern blot 分析显示的 DNA 条带 位置发生改变。人群存在的遗传差异性叫多态性。这种突变可能本身就是致病变异,也有 可能与致病变异紧密连锁。遗传疾病如镰刀性贫血症、囊性纤维病变、Huntington chorea 可以用 RFLP 分析鉴定

选择性终止复制能测定DNA序列 DNA序列测定技术也极大地促进了DNA结构和功能的研究。DNA序列测定的关键是 产生一组DNA片段,其长度取决于DNA序列最后一个核苷酸。Frederick Sanger及其同事 建立了Sanger双脱氧方法(即选择性终止复制)测定DNA序列的技术。与其他技术相比, Sanger双脱氧法简便,能够同时进行四种终止反应。在这些混合物中,DNA聚合酶用来制 造DNA单链模板的互补DNA序列。合成引物是化学合成的DNA片段,与模板分子的序列 已知区互补。除了四种脱氧核苷三磷酸(同位素标记)外,每个反应混合物都含有少量的一 种2,3双脱氧核苷三磷酸类似物,各个混合物所含的类似物不同。 DNA to be sequenced 3'—GAATTCGCTAATGG- 5'—CTTAA Primer DNA polymerase I Labeled dATP TTP dCTP,dGTP Dideoxy analog of dATP 3-GAATTCGCTAATGC 5-CTTAAGCGATTA + 3-GAATTCGCTAATGG 5'—CTTAAGCGA New DNA strands are separated and subjected to electrophoresis Figure 5-4 2',3'-Dideoxy analog 图5.4链终止法测定DNA序列的策略。一种2'3'-双脱氧核苷三磷酸加入DNA聚合酶促反 应混合物产生一组DNA片段,其3末端就是这种2'3-双脱氧核苷三磷酸。例如ddATP(红 色)产生的DNA片段,其末端都是ddA。由于缺乏3'OH,这种链不能继续延伸。 由于核苷类似物3'末端缺乏OH,这种类似物的参入能够阻止生长链进一步延长。双脱 氧类似物浓度要足够低,使生长链偶尔参入一个双脱氧核苷酸导致DNA合成终止。DNA 聚合酶有时参入正常的核苷酸,有时参入双脱氧核苷酸导致反应终止。例如,反应混合物含 有ddATP产生的合成DNA链长度不同,但是3'-端都是ddA(图5.4)。ddA参与的位点就是 待测核酸链的T。因此根据合成核酸链长度就能确定待测核酸链T的位置。一套(四种终止 混合物)双脱氧终止反应后,上样到凝胶的四个孔中电泳分离,从放射性自显影图谱能读出 待测核酸序列。 荧光检测的灵敏度很高,能够替代放射性同位素。每个双脱氧核苷酸连接一个荧光标签, 这样每种链终止反应就带有一种特异的荧光标签(例如蓝色荧光表示ddA,红色荧光表示 dC)。此时能够将四种终止反应混合物混在一起,进行聚合酶链终止反应。产物在一个上样 孔中进行凝胶电泳分离。条带泳动的先后次序表示DNA链的长度,条带的荧光颜色表示 DNA链的终止核苷酸,由此测定DNA链的核酸序列(图5.5)。这种方式能够测定长度达 到550核苷酸的DNA序列。荧光检测更好,这种方法不用放射性同位素,易于自动化检测。 实际上,现代DNA测序仪器采用这种方法每天能够测定的DNA序列超过1O0万碱基
选择性终止复制能测定 DNA 序列 DNA 序列测定技术也极大地促进了 DNA 结构和功能的研究。DNA 序列测定的关键是 产生一组 DNA 片段,其长度取决于 DNA 序列最后一个核苷酸。Frederick Sanger 及其同事 建立了 Sanger 双脱氧方法(即选择性终止复制)测定 DNA 序列的技术。与其他技术相比, Sanger 双脱氧法简便,能够同时进行四种终止反应。在这些混合物中,DNA 聚合酶用来制 造 DNA 单链模板的互补 DNA 序列。合成引物是化学合成的 DNA 片段,与模板分子的序列 已知区互补。除了四种脱氧核苷三磷酸(同位素标记)外,每个反应混合物都含有少量的一 种 2', 3'-双脱氧核苷三磷酸类似物,各个混合物所含的类似物不同。 图 5.4 链终止法测定 DNA 序列的策略。一种 2'3'-双脱氧核苷三磷酸加入 DNA 聚合酶促反 应混合物产生一组 DNA 片段,其 3'-末端就是这种 2'3'-双脱氧核苷三磷酸。例如 ddATP(红 色)产生的 DNA 片段,其末端都是 ddA。由于缺乏 3'-OH,这种链不能继续延伸。 由于核苷类似物 3'-末端缺乏 OH,这种类似物的参入能够阻止生长链进一步延长。双脱 氧类似物浓度要足够低,使生长链偶尔参入一个双脱氧核苷酸导致 DNA 合成终止。DNA 聚合酶有时参入正常的核苷酸,有时参入双脱氧核苷酸导致反应终止。例如,反应混合物含 有 ddATP 产生的合成 DNA 链长度不同,但是 3'-端都是 ddA(图 5.4)。ddA 参与的位点就是 待测核酸链的 T。因此根据合成核酸链长度就能确定待测核酸链 T 的位置。一套(四种终止 混合物)双脱氧终止反应后,上样到凝胶的四个孔中电泳分离,从放射性自显影图谱能读出 待测核酸序列。 荧光检测的灵敏度很高,能够替代放射性同位素。每个双脱氧核苷酸连接一个荧光标签, 这样每种链终止反应就带有一种特异的荧光标签(例如蓝色荧光表示 ddA, 红色荧光表示 ddC)。此时能够将四种终止反应混合物混在一起,进行聚合酶链终止反应。产物在一个上样 孔中进行凝胶电泳分离。条带泳动的先后次序表示 DNA 链的长度,条带的荧光颜色表示 DNA 链的终止核苷酸,由此测定 DNA 链的核酸序列(图 5.5)。这种方式能够测定长度达 到 550 核苷酸的 DNA 序列。荧光检测更好,这种方法不用放射性同位素,易于自动化检测。 实际上,现代 DNA 测序仪器采用这种方法每天能够测定的 DNA 序列超过 100 万碱基