
第十四章信号传导途径 0 GTP "ON" position position GTP 生物系统的信号传导通路是分子开关的接通和关闭。如同计算机芯片(左边),处于“接通” 状态就能传递信息。这些通路的共同点一些蛋白质(包括G蛋白,右边),与GTP结合的 能够传递信号,与GDP结合就处于沉默状态。 细胞对环境特异化学物质应答程度很高。在感知环境有这种化学物质是,细胞的应答也 许改变细胞的代谢,也许改变细胞的基因表达模式。在多细胞生物,这些化学信号是协调生 物应答的关键(图14.1)。刺激生理应答的三个分子信号的实例分别是肾上腺素、胰岛素、 和表皮生长因子。当生物个体受到威胁,处于肾脏上面的肾上腺释放肾上腺素。肾上腺素能 促进能源转化,改进心脏功能。进食后,胰岛的阝-细胞释放胰岛素。胰岛素刺激机体从血 液摄取葡萄糖,产生其它生理变化。应答损伤时,机体释放表皮生长因子刺激特异细胞生长 和分裂。上述的各种情况,细胞都是接受到一种信息,即环境中某种分子的浓度已经超过阙 值,启动生物的一系列事件将“环境中这种分子的浓度超过阈值”的信息转化成生理应答, 这个过程叫信号传导(signal transduction)。 Epinephrine Insulin Epidermal growth factor(EGF) B-Adrenergic receptor Insulin receptor EGF receptor Energy-store Increased glucose Expression of mobilization uptake growth-promoting genes 图14.1三种信号传导途径。信号分子与受体结合启动相应的生物过程,使机体产生重要的 生理应答。 根据信号传导途径的组分及其分支情况,对信号传导途径进行定义。这些信号传导途径 非常复杂,易于混淆。然而,很多信号传导途径所采用的策略和分子的种类又不乏有很多共 性。这些共同特性所遵循的原则能揭示信号传导途径的逻辑。由于信号传导途径影响本书其 余章节将要介绍的代谢途径的各个方面,我们在此介绍信号传导途径的这些原理
第十四章 信号传导途径 生物系统的信号传导通路是分子开关的接通和关闭。如同计算机芯片(左边),处于“接通” 状态就能传递信息。这些通路的共同点一些蛋白质(包括 G 蛋白,右边),与 GTP 结合的 能够传递信号,与 GDP 结合就处于沉默状态。 细胞对环境特异化学物质应答程度很高。在感知环境有这种化学物质是,细胞的应答也 许改变细胞的代谢,也许改变细胞的基因表达模式。在多细胞生物,这些化学信号是协调生 物应答的关键(图 14.1)。刺激生理应答的三个分子信号的实例分别是肾上腺素、胰岛素、 和表皮生长因子。当生物个体受到威胁,处于肾脏上面的肾上腺释放肾上腺素。肾上腺素能 促进能源转化,改进心脏功能。进食后,胰岛的 细胞释放胰岛素。胰岛素刺激机体从血 液摄取葡萄糖,产生其它生理变化。应答损伤时,机体释放表皮生长因子刺激特异细胞生长 和分裂。上述的各种情况,细胞都是接受到一种信息,即环境中某种分子的浓度已经超过阈 值,启动生物的一系列事件将“环境中这种分子的浓度超过阈值”的信息转化成生理应答, 这个过程叫信号传导(signal transduction)。 图 14.1 三种信号传导途径。信号分子与受体结合启动相应的生物过程,使机体产生重要的 生理应答。 根据信号传导途径的组分及其分支情况,对信号传导途径进行定义。这些信号传导途径 非常复杂,易于混淆。然而,很多信号传导途径所采用的策略和分子的种类又不乏有很多共 性。这些共同特性所遵循的原则能揭示信号传导途径的逻辑。由于信号传导途径影响本书其 余章节将要介绍的代谢途径的各个方面,我们在此介绍信号传导途径的这些原理

信号传导以来分子通路 Signal V Reception Amplification Transduction Response(s) 图142信号传导原则。一个细胞组分首先与环境信号结合。这个细胞组分常常是细胞表面 受体。然后将信号已经到达的信息转化成其他化学形式,或转导。转导过程常常有很多步骤 构成。在发生最后应答前,这些步骤常常依次扩增反馈途径调节整个信号途径。 信号传导途径可以看作是一种分子通路(图14.2)。所有这些分子通路都含有一些关键 步骤: 1.第一信使的释放。一个刺激(如损伤或消化食物)启动信号分子释放。这些信号分子叫 第一信使。 2.第一信使的接收。大多数信号分子不进入细胞。细胞表面的蛋白质充当受体,能够与信 号分子结合,然后将它们已经与信号分子结合的“信息”传递到细胞内。受体跨过细胞 膜,因此既有胞内组分又有胞外组分。特异识别信号分子(也称为陪体)的结合位点就 在胞外。这种结合位点与酶的活性位点相似,只是没有在此处发生催化作用。配体与受 体之间的相互作用改变了受体蛋白的三级或四级结构,包括受体蛋白在细胞内组分的空 间结构。 3.在细胞内用第二信使传递信息。被称为第二信使的另外一些小分子用来接着将受体-配 体复合物的信息向下传递。第二信使是胞内分子,其浓度是应答环境信号的。这些浓度 变化构成分子通路的下一环节。特别重要的第二信使是cAMP和cGMP,Ca+,肌醇 -1,3,5-三磷酸(IP3),和二酰基甘油(DAG,图14.3)。 24 H20 OH OPO,2- +o;Po OH H20 <AMP,(GMP Calcium ion Inositol 1,4,5-trisphosphate (IPa) OH Diacylglycerol(DAG 图14.3常用的第二信使。第二信使是胞内分子,其浓度在应答环境信号时发生改变,在细 胞内传送信息
信号传导以来分子通路 图 14.2 信号传导原则。一个细胞组分首先与环境信号结合。这个细胞组分常常是细胞表面 受体。然后将信号已经到达的信息转化成其他化学形式,或转导。转导过程常常有很多步骤 构成。在发生最后应答前,这些步骤常常依次扩增反馈途径调节整个信号途径。 信号传导途径可以看作是一种分子通路(图 14.2)。所有这些分子通路都含有一些关键 步骤: 1. 第一信使的释放。一个刺激(如损伤或消化食物)启动信号分子释放。这些信号分子叫 第一信使。 2. 第一信使的接收。大多数信号分子不进入细胞。细胞表面的蛋白质充当受体,能够与信 号分子结合,然后将它们已经与信号分子结合的“信息”传递到细胞内。受体跨过细胞 膜,因此既有胞内组分又有胞外组分。特异识别信号分子(也称为陪体)的结合位点就 在胞外。这种结合位点与酶的活性位点相似,只是没有在此处发生催化作用。配体与受 体之间的相互作用改变了受体蛋白的三级或四级结构,包括受体蛋白在细胞内组分的空 间结构。 3. 在细胞内用第二信使传递信息。被称为第二信使的另外一些小分子用来接着将受体-配 体复合物的信息向下传递。第二信使是胞内分子,其浓度是应答环境信号的。这些浓度 变化构成分子通路的下一环节。特别重要的第二信使是 cAMP 和 cGMP,Ca 2+ , 肌醇 -1,3,5-三磷酸(IP3),和二酰基甘油(DAG; 图 14.3)。 图 14.3 常用的第二信使。第二信使是胞内分子,其浓度在应答环境信号时发生改变,在细 胞内传送信息

采用第二信使产生了几种结果。首先,在产生第二信使之前信号可能已被放大。信号分 子与受体结合可能只活化了少量受体分子,但是每个活化的受体分子能产生很多第二信使。 因此,低浓度的环境信号分子,即使少到只有一个信号分子,也能产生大量的胞内信号和应 答。其次,第二信使分子能自由扩散到细胞其他区域,因此能够影响整个细胞。第三,在多 个信号途径采用相同的第二信使既造就了一些机会,也产生了一些问题。从几个信号途径输 入信号(常常称为Cross-talk),会玫变共同第二信使的浓度。与单独途径作用相比,Cross talk 可以精细调节细胞活性。但是,不当的cross talk导致的第二信使浓度改变可能被细胞误读。 4.直接改变生理应答的效应器活化。信号途径的基本效应是活化(或抑制)泵、酶、和直 接控制代谢途径的基因转录因子,基因活化,以及神经信号传递。 5.信号终止。在细胞已经完成了对信号的应答之后,必须终止信号途径或者不再对新的信 号实施应答。不能适当终止信号应答的细胞会导致非常严重的后果。我们将看到,很多 肿瘤细胞与它们不能适当地终止信号途径有关,尤其是控制细胞生长的生物过程。 本章我们介绍图14.1提到的三种信号途径的各个组分。在介绍过程中,我们会看到信 号转导蛋白的几类接头(adaptor)结构域。这些结构域通常能识别特定分子,有助于将信息 从一种蛋白质传递到另一种蛋白质。这三种信号传递途径的组分也出现于其他信号传导途 径。记住,这三个信号传导途径也是很多其他信号传导途径的代表。 14.1异源三聚体G蛋白传递信号、重置自身 B-肾上腺素受体(B-AR)与肾上腺素配体结合,起始肾上腺素信号途径。B-肾上腺素 受体属于细胞表面受体家族的成员。这个家族是TTM受体家族(seven transmembrane-helix receptors)。该家族成员负贡传递不同信号启动的信息。这些信号包括激素、神经递质、气味 (odorants)、味觉物质、甚至质子(表l4.1)。已经知道的7TM受体家族成员有几千种。约 有50%的治疗药物靶向这类受体。该家族蛋白有7个α-螺旋跨过膜脂质双层。由于是单一 多肽链如同蛇形跨膜7次,也有人称这类受体为“蛇形卷曲受体”(serpentine receptors)(图 14.4A)。最先被确定三维结构的7TM受体是视紫质(图14.4B),在视觉系统其关键作用。 其他很多TTM受体家族成员的结构域视紫质蛋白结构很相似,有些7TM受体家族成员保 外结构域较大。 表14.17TM受体介导的生物功能 TABLE 14.I Biological functions mediated by 7TM receptors ◆Hormone action Hormone secretion Neurotransmission Chemotaxis ·Exocytosis Control of blood pressure ·Embryogenesis Cell growth and differentiation Development ●Smell ●Taste 。Vision .Viral infection Source:After J.S.Gutkind,J.Biol.Chem.273(1998): 1839-1842
采用第二信使产生了几种结果。首先,在产生第二信使之前信号可能已被放大。信号分 子与受体结合可能只活化了少量受体分子,但是每个活化的受体分子能产生很多第二信使。 因此,低浓度的环境信号分子,即使少到只有一个信号分子,也能产生大量的胞内信号和应 答。其次,第二信使分子能自由扩散到细胞其他区域,因此能够影响整个细胞。第三,在多 个信号途径采用相同的第二信使既造就了一些机会,也产生了一些问题。从几个信号途径输 入信号(常常称为 Cross-talk),会改变共同第二信使的浓度。与单独途径作用相比,Cross talk 可以精细调节细胞活性。但是,不当的 cross talk 导致的第二信使浓度改变可能被细胞误读。 4. 直接改变生理应答的效应器活化。信号途径的基本效应是活化(或抑制)泵、酶、和直 接控制代谢途径的基因转录因子,基因活化,以及神经信号传递。 5. 信号终止。在细胞已经完成了对信号的应答之后,必须终止信号途径或者不再对新的信 号实施应答。不能适当终止信号应答的细胞会导致非常严重的后果。我们将看到,很多 肿瘤细胞与它们不能适当地终止信号途径有关,尤其是控制细胞生长的生物过程。 本章我们介绍图 14.1 提到的三种信号途径的各个组分。在介绍过程中,我们会看到信 号转导蛋白的几类接头(adaptor)结构域。这些结构域通常能识别特定分子,有助于将信息 从一种蛋白质传递到另一种蛋白质。这三种信号传递途径的组分也出现于其他信号传导途 径。记住,这三个信号传导途径也是很多其他信号传导途径的代表。 14.1 异源三聚体 G 蛋白传递信号、重置自身 肾上腺素受体(AR)与肾上腺素配体结合,起始肾上腺素信号途径。肾上腺素 受体属于细胞表面受体家族的成员。这个家族是 7TM 受体家族(seven transmembrane-helix receptors)。该家族成员负责传递不同信号启动的信息。这些信号包括激素、神经递质、气味 (odorants)、味觉物质、甚至质子(表 14.1)。已经知道的 7TM 受体家族成员有几千种。约 有 50%的治疗药物靶向这类受体。该家族蛋白有 7 个 螺旋跨过膜脂质双层。由于是单一 多肽链如同蛇形跨膜 7 次,也有人称这类受体为“蛇形卷曲受体”(serpentine receptors)(图 14.4A)。最先被确定三维结构的 7TM 受体是视紫质(图 14.4B),在视觉系统其关键作用。 其他很多 7TM 受体家族成员的结构域视紫质蛋白结构很相似,有些 7TM 受体家族成员保 外结构域较大。 表 14.1 7TM 受体介导的生物功能
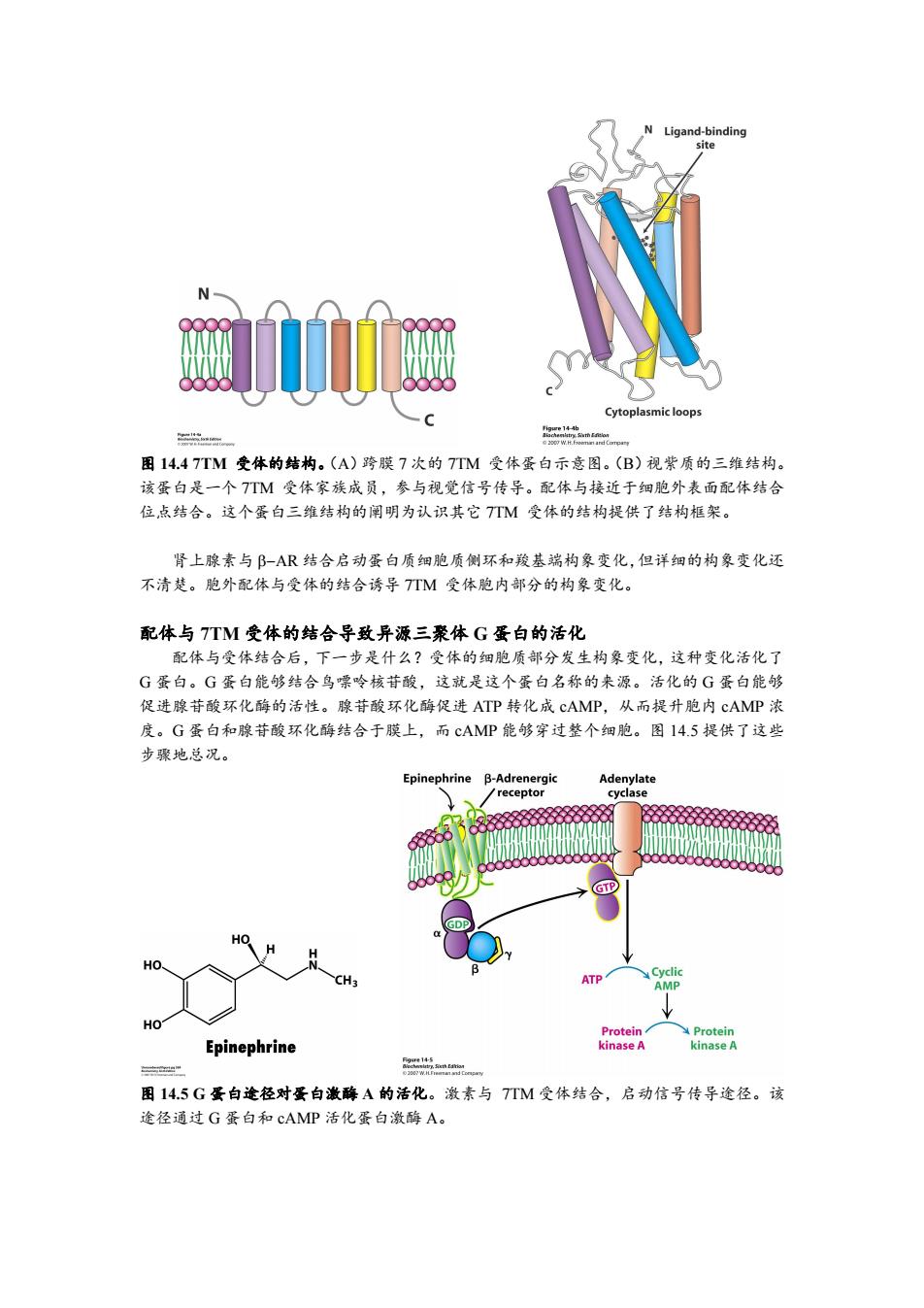
Ligand-binding site Cytoplasmic loops 图14.47TM受体的结构。(A)跨膜7次的7TM受体蛋白示意图。(B)视紫质的三维结构。 该蛋白是一个TTM受体家族成员,参与视觉信号传导。配体与接近于细胞外表面配体结合 位,点结合。这个蛋白三维结构的阐明为认识其它7TM受体的结构提供了结构框架。 肾上腺素与阝-AR结合启动蛋白质细胞质侧环和羧基端构象变化,但详细的构象变化还 不清楚。胞外配体与受体的结合诱导TTM受体胞内部分的构象变化。 配体与7TM受体的结合导致异源三聚体G蛋白的活化 配体与受体结合后,下一步是什么?受体的细胞质部分发生构象变化,这种变化活化了 G蛋白。G蛋白能够结合鸟嘌呤核苷酸,这就是这个蛋白名称的来源。活化的G蛋白能够 促进腺苷酸环化酶的活性。腺苷酸环化酶促进ATP转化成cAMP,从而提升胞内cAMP浓 度。G蛋白和腺苷酸环化酶结合于膜上,而cAMP能够穿过整个细胞。图14.5提供了这些 步骤地总况。 Epinephrine B-Adrenergic Adenylate receptor cyclase GTP 0 HO HO. CH3 ATP Cyclic AMP HO Protein Protein Epinephrine kinase A kinase A " 图14.5G蛋白途径对蛋白激酶A的活化。激素与7TM受体结合,启动信号传导途径。该 途径通过G蛋白和cAMP活化蛋白激酶A
图 14.4 7TM 受体的结构。(A)跨膜 7 次的 7TM 受体蛋白示意图。(B)视紫质的三维结构。 该蛋白是一个 7TM 受体家族成员,参与视觉信号传导。配体与接近于细胞外表面配体结合 位点结合。这个蛋白三维结构的阐明为认识其它 7TM 受体的结构提供了结构框架。 肾上腺素与 AR 结合启动蛋白质细胞质侧环和羧基端构象变化,但详细的构象变化还 不清楚。胞外配体与受体的结合诱导 7TM 受体胞内部分的构象变化。 配体与 7TM 受体的结合导致异源三聚体 G 蛋白的活化 配体与受体结合后,下一步是什么?受体的细胞质部分发生构象变化,这种变化活化了 G 蛋白。G 蛋白能够结合鸟嘌呤核苷酸,这就是这个蛋白名称的来源。活化的 G 蛋白能够 促进腺苷酸环化酶的活性。腺苷酸环化酶促进 ATP 转化成 cAMP,从而提升胞内 cAMP 浓 度。G 蛋白和腺苷酸环化酶结合于膜上,而 cAMP 能够穿过整个细胞。图 14.5 提供了这些 步骤地总况。 图 14.5 G 蛋白途径对蛋白激酶 A 的活化。激素与 7TM 受体结合,启动信号传导途径。该 途径通过 G 蛋白和 cAMP 活化蛋白激酶 A
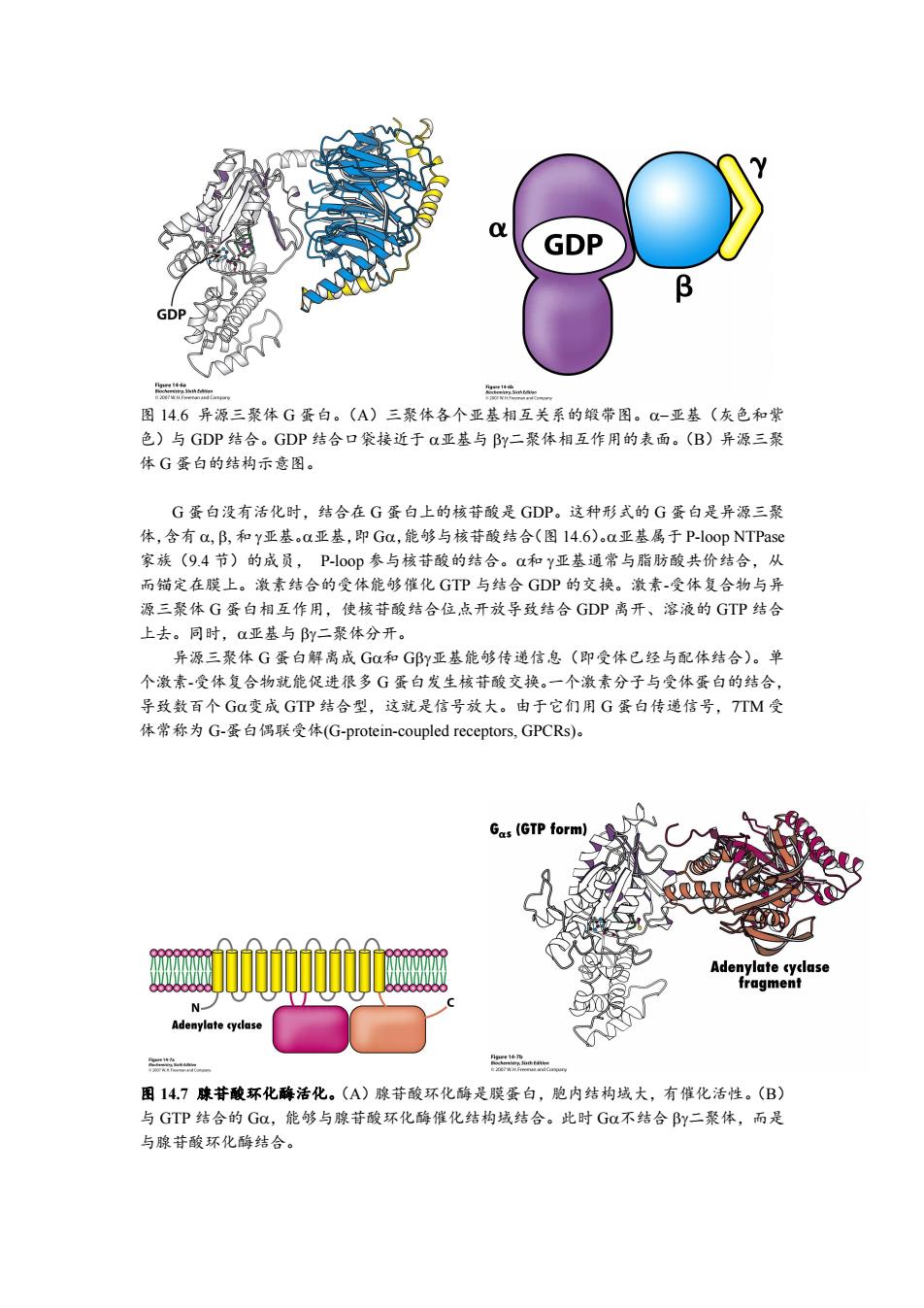
GDP B GD 图14.6异源三聚体G蛋白。(A)三聚体各个亚基相互关系的缎带图。-亚基(灰色和紫 色)与GDP结合。GDP结合口袋接近于a亚基与By二聚体相互作用的表面。(B)异源三聚 体G蛋白的结构示意图。 G蛋白没有活化时,结合在G蛋白上的核苷酸是GDP。这种形式的G蛋白是异源三聚 体,含有a,B,和Y亚基。a亚基,即Ga,能够与核苷酸结合(图14.6)a亚基属于P--loop NTPase 家族(9.4节)的成员,P-00p参与核苷酸的结合。Q和Y亚基通常与脂肪酸共价结合,从 而锚定在膜上。激素结合的受体能够催化GTP与结合GDP的交换。激素-受体复合物与异 源三聚体G蛋白相互作用,使核苷酸结合位,点开放导致结合GDP离开、溶液的GTP结合 上去。同时,亚基与By二聚体分开。 异源三聚体G蛋白解离成G和G邱y亚基能够传递信息(即受体已经与配体结合)。单 个激素-受体复合物就能促进很多G蛋白发生核苷酸交换。一个激素分子与受体蛋白的结合, 导致数百个Ga变成GTP结合型,这就是信号放大。由于它们用G蛋白传递信号,TTM受 体常称为G-蛋白偶联受体(G-protein-coupled receptors,GPCRs)。 Gas(GTP form) Adenylate cyclase fragment N- Adenylate cyclase 一 二- 图14.7腺苷酸环化酶活化。(A)腺苷酸环化酶是膜蛋白,胞内结构域大,有催化活性。(B) 与GTP结合的GC,能够与腺苷酸环化酶催化结构域结合。此时G不结合By二聚体,而是 与腺苷酸环化酶结合
图 14.6 异源三聚体 G 蛋白。(A)三聚体各个亚基相互关系的缎带图。亚基(灰色和紫 色)与 GDP 结合。GDP 结合口袋接近于 亚基与 二聚体相互作用的表面。(B)异源三聚 体 G 蛋白的结构示意图。 G 蛋白没有活化时,结合在 G 蛋白上的核苷酸是 GDP。这种形式的 G 蛋白是异源三聚 体,含有 和 亚基。亚基,即 G,能够与核苷酸结合(图 14.6)。亚基属于 P-loop NTPase 家族(9.4 节)的成员, P-loop 参与核苷酸的结合。和 亚基通常与脂肪酸共价结合,从 而锚定在膜上。激素结合的受体能够催化 GTP 与结合 GDP 的交换。激素-受体复合物与异 源三聚体 G 蛋白相互作用,使核苷酸结合位点开放导致结合 GDP 离开、溶液的 GTP 结合 上去。同时,亚基与 二聚体分开。 异源三聚体 G 蛋白解离成 G和 G亚基能够传递信息(即受体已经与配体结合)。单 个激素-受体复合物就能促进很多 G 蛋白发生核苷酸交换。一个激素分子与受体蛋白的结合, 导致数百个 G变成 GTP 结合型,这就是信号放大。由于它们用 G 蛋白传递信号,7TM 受 体常称为 G-蛋白偶联受体(G-protein-coupled receptors, GPCRs)。 图 14.7 腺苷酸环化酶活化。(A)腺苷酸环化酶是膜蛋白,胞内结构域大,有催化活性。(B) 与 GTP 结合的 G,能够与腺苷酸环化酶催化结构域结合。此时 G不结合 二聚体,而是 与腺苷酸环化酶结合