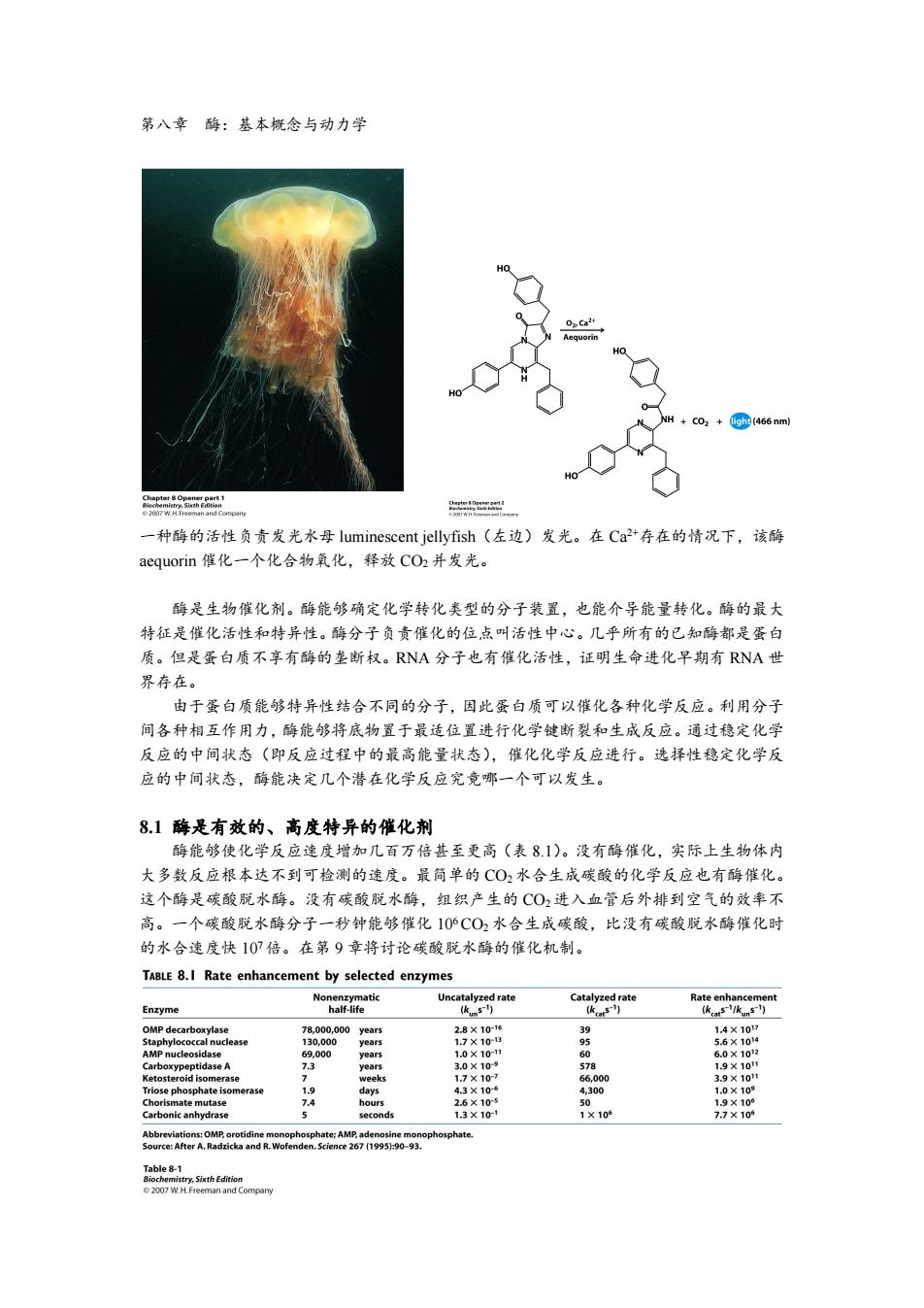
第八章酶:基本概念与动力学 0Ca24 equorin +CO2 (466 nm) 9 一种酶的活性负责发光水母luminescent jellyfish(左边)发光。在Ca2+存在的情况下,该酶 aequorin催化一个化合物氧化,释放CO2并发光。 酶是生物催化剂。酶能够确定化学转化类型的分子装置,也能介导能量转化。酶的最大 特征是催化活性和特异性。酶分子负贡催化的位点叫活性中心。几乎所有的已知酶都是蛋白 质。但是蛋白质不享有酶的垄断权。RNA分子也有催化活性,证明生命进化早期有RNA世 界存在。 由于蛋白质能够特异性结合不同的分子,因此蛋白质可以催化各种化学反应。利用分子 间各种相互作用力,酶能够将底物置于最适位置进行化学键断裂和生成反应。通过稳定化学 反应的中间状态(即反应过程中的最高能量状态),催化化学反应进行。选择性稳定化学反 应的中间状态,酶能决定几个潜在化学反应究竟哪一个可以发生。 8.1酶是有效的、高度特异的催化剂 酶能够使化学反应速度增加几百万倍甚至更高(表8.1)。没有酶催化,实际上生物体内 大多数反应根本达不到可检测的速度。最简单的CO2水合生成碳酸的化学反应也有酶催化。 这个酶是碳酸脱水酶。没有碳酸脱水酶,组织产生的CO2进入血管后外排到空气的效率不 高。一个碳酸脱水酶分子一秒钟能够催化106C02水合生成碳酸,比没有碳酸脱水酶催化时 的水合速度快107倍。在第9章将讨论碳酸脱水酶的催化机制。 TABLE 8.I Rate enhancement by selected enzymes Nonenzymatic Uncatalyzed rate Catalyzed rate Rate enhancement Enzyme half-life (km51) (ka5) (kcats-/kuns-) OMP decarboxylase 78,000,000year5 2.8×1016 39 1.4×107 Staphylococcal nuclease 130,000 years 1.7×10-13 9 5.6×1014 AMP nucleosidase 69.000 years 1.0×10-11 60 6.0×1012 Carboxypeptidase A 73 years 3.0×10- 578 1.9×10t Ketosteroid isomerase 2 weeks 1.7×10-7 66,000 3.9×101 Triose phosphate isomerase 1.9 days 4.3×106 4,300 1.0×109 Chorismate mutase 7.4 hours 2.6×10-5 50 1.9×105 Carbonic anhydrase seconds 13×10-1 1×10 7.7×10 Abbreviations:OMP,orotidine monophosphate:AMP,adenosine monophosphate. Source:After A.Radzicka and R.Wofenden.Science 267(1995):90-93. Table8-1 Biochemistry,Sixth Edition 2007 W.H.Freeman and Compary
第八章 酶:基本概念与动力学 一种酶的活性负责发光水母 luminescent jellyfish(左边)发光。在 Ca 2+存在的情况下,该酶 aequorin 催化一个化合物氧化,释放 CO2并发光。 酶是生物催化剂。酶能够确定化学转化类型的分子装置,也能介导能量转化。酶的最大 特征是催化活性和特异性。酶分子负责催化的位点叫活性中心。几乎所有的已知酶都是蛋白 质。但是蛋白质不享有酶的垄断权。RNA 分子也有催化活性,证明生命进化早期有 RNA 世 界存在。 由于蛋白质能够特异性结合不同的分子,因此蛋白质可以催化各种化学反应。利用分子 间各种相互作用力,酶能够将底物置于最适位置进行化学键断裂和生成反应。通过稳定化学 反应的中间状态(即反应过程中的最高能量状态),催化化学反应进行。选择性稳定化学反 应的中间状态,酶能决定几个潜在化学反应究竟哪一个可以发生。 8.1 酶是有效的、高度特异的催化剂 酶能够使化学反应速度增加几百万倍甚至更高(表 8.1)。没有酶催化,实际上生物体内 大多数反应根本达不到可检测的速度。最简单的 CO2水合生成碳酸的化学反应也有酶催化。 这个酶是碳酸脱水酶。没有碳酸脱水酶,组织产生的 CO2 进入血管后外排到空气的效率不 高。一个碳酸脱水酶分子一秒钟能够催化 10 6CO2水合生成碳酸,比没有碳酸脱水酶催化时 的水合速度快 10 7倍。在第 9 章将讨论碳酸脱水酶的催化机制
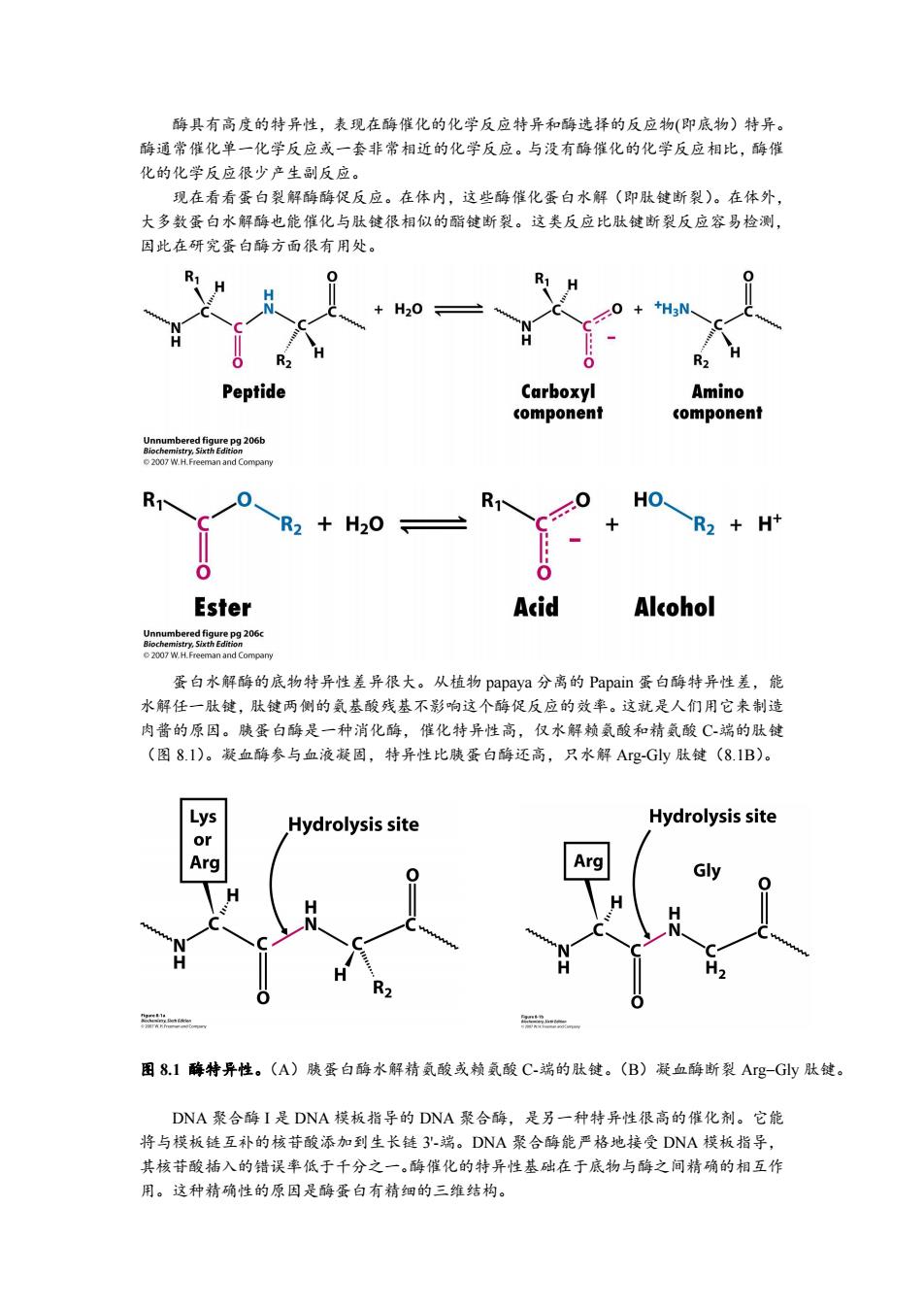
酶具有高度的特异性,表现在酶催化的化学反应特异和酶选择的反应物(即底物)特异。 酶通常催化单一化学反应或一套非常相近的化学反应。与没有酶催化的化学反应相比,酶催 化的化学反应很少产生副反应。 现在看看蛋白裂解酶酶促反应。在体内,这些酶催化蛋白水解(即肽键断裂)。在体外, 大多数蛋白水解酶也能催化与肽键很相似的酯键断裂。这类反应比肽键断裂反应容易检测, 因此在研究蛋白酶方面很有用处。 H20 Peptide Carboxyl Amino component component Unnumbered figure pg 206b 2007 W.H.Freeman and Compary R HO R2 +H20 R2 H+ Ester Acid Alcohol cm06 2007 W.H.Freeman and Company 蛋白水解酶的底物特异性差异很大。从植物papaya分离的Papain蛋白酶特异性差,能 水解任一肽键,肽键两侧的氨基酸残基不影响这个酶促反应的效率。这就是人们用它来制造 肉酱的原因。胰蛋白酶是一种消化酶,催化特异性高,仅水解赖氨酸和精氨酸C端的肽键 (图8.I)。凝血酶参与血液凝固,特异性比胰蛋白酶还高,只水解Ag-Gy肽键(8.1B)。 Hydrolysis site Hydrolysis site Arg 2n 图8.1酶特异性。(A)胰蛋白酶水解精氨酸或赖氨酸C-端的肽键。(B)凝血酶断裂Ag-Gly肽键。 DNA聚合酶I是DNA模板指导的DNA聚合酶,是另一种特异性很高的催化剂。它能 将与模板链互补的核苷酸添加到生长链3'-端。DNA聚合酶能严格地接受DNA模板指导, 其核苷酸插入的错误率低于千分之一。酶催化的特异性基础在于底物与酶之间精确的相互作 用。这种精确性的原因是酶蛋白有精细的三维结构
酶具有高度的特异性,表现在酶催化的化学反应特异和酶选择的反应物(即底物)特异。 酶通常催化单一化学反应或一套非常相近的化学反应。与没有酶催化的化学反应相比,酶催 化的化学反应很少产生副反应。 现在看看蛋白裂解酶酶促反应。在体内,这些酶催化蛋白水解(即肽键断裂)。在体外, 大多数蛋白水解酶也能催化与肽键很相似的酯键断裂。这类反应比肽键断裂反应容易检测, 因此在研究蛋白酶方面很有用处。 蛋白水解酶的底物特异性差异很大。从植物 papaya 分离的 Papain 蛋白酶特异性差,能 水解任一肽键,肽键两侧的氨基酸残基不影响这个酶促反应的效率。这就是人们用它来制造 肉酱的原因。胰蛋白酶是一种消化酶,催化特异性高,仅水解赖氨酸和精氨酸 C-端的肽键 (图 8.1)。凝血酶参与血液凝固,特异性比胰蛋白酶还高,只水解 Arg-Gly 肽键(8.1B)。 图 8.1 酶特异性。(A)胰蛋白酶水解精氨酸或赖氨酸 C-端的肽键。(B)凝血酶断裂 ArgGly 肽键。 DNA 聚合酶 I 是 DNA 模板指导的 DNA 聚合酶,是另一种特异性很高的催化剂。它能 将与模板链互补的核苷酸添加到生长链 3'-端。DNA 聚合酶能严格地接受 DNA 模板指导, 其核苷酸插入的错误率低于千分之一。酶催化的特异性基础在于底物与酶之间精确的相互作 用。这种精确性的原因是酶蛋白有精细的三维结构

很多酶其活性需要辅助因子 很多酶的催化活性依赖于被称为辅助因子的小分子(虽然辅助因子在不同酶的作用并不 相同)。通常这些辅助因子能够执行20种氨基酸不能执行的化学反应。没有辅助因子的酶叫 脱辅基酶(apoenzyme):全酶(holoenzyme)是具有催化活性的酶。 Apoenzyme cofactor =holoenzyme 辅助因子可以分为两类:(1)金属和(2)小的有机分子(称为辅酶)(表8.2)。辅酶 常常是维生素的衍生物,与酶结合有的松懈,有的紧密。紧密结合的辅酶叫辅基,结合松弛 的辅酶像协同底物(cosubstrate)。这些辅酶与酶的结合与解离类似酶促反应的第底物和产物。 不同的酶利用相同的辅酶,因此辅酶与正常的底物是不同的。利用同一辅酶的酶,虽然酶促 反应不同,但是这些酶催化的机理类似。在第9章,我们将讨论金属离子对酶活性的影响。 在整个生物化学教科书,我们会经常介绍辅酶与相应的酶蛋白之间的互作。后面我们会详细 介绍维生素知识。 TABLE 8.2 Enzyme cofactors Cofactor Enzyme Coenzyme Thiamine pyrophosphate Pyruvate dehydrogenase Flavin adenine nucleotide Monoamine oxidase Nicotinamide adenine dinucleotide Lactate dehydrogenase Pyridoxal phosphate Glycogen phosphorylase Coenzyme A(CoA) Acetyl CoA carboxylase Biotin Pyruvate carboxylase 5'-Deoxyadenosyl cobalamin Methylmalonyl mutase Tetrahydrofolate Thymidylate synthase Metal Zn2+ Carbonic anhydrase Zn2+ Carboxypeptidase Mg2+ EcoRV Mg2+ Hexokinase Ni2+ Urease Mo Nitrate reductase Se Glutathione peroxidase Mn Superoxide dismutase K+ Propionyl CoA carboxylase Table 8-2 try,Sixth Edition 2007 W.H.Freeman and Company 酶能够将一种形式的能量转化成另一种形式 很多生化反应中,反应物的能量能够高效转化成其他能量形式。例如,光合成反应能够 将光能转化成化学能。线粒体能够将食物小分子的自由能转化成离子梯度自由能,再转化成 ATP(一种通用的能量分子)。酶能够以很多方式利用ATP的化学键能。例如,肌球蛋白 (myosin)能够将ATP的能量转化成肌肉收缩的机械能(34章)膜上的泵是一种酶,能够 利用ATP的能量驱动分子和离子跨膜运输(第13章)。化学物质和离子不对称分布所形成 的浓度梯度也是一种能量形式,有不同的用途,如发射神经脉冲。 科学家正在阐明这些能量转导酶作用的分子机制。后面章节我们将介绍不同步骤(结合、 化学转化、和释放)是如何单向循环导致能量从一种形式转化成另一种形式的机制
很多酶其活性需要辅助因子 很多酶的催化活性依赖于被称为辅助因子的小分子(虽然辅助因子在不同酶的作用并不 相同)。通常这些辅助因子能够执行 20 种氨基酸不能执行的化学反应。没有辅助因子的酶叫 脱辅基酶(apoenzyme);全酶(holoenzyme)是具有催化活性的酶。 Apoenzyme + cofactor == holoenzyme 辅助因子可以分为两类:(1)金属和 (2)小的有机分子(称为辅酶)(表 8.2)。辅酶 常常是维生素的衍生物,与酶结合有的松懈,有的紧密。紧密结合的辅酶叫辅基,结合松弛 的辅酶像协同底物(cosubstrate)。这些辅酶与酶的结合与解离类似酶促反应的第底物和产物。 不同的酶利用相同的辅酶,因此辅酶与正常的底物是不同的。利用同一辅酶的酶,虽然酶促 反应不同,但是这些酶催化的机理类似。在第 9 章,我们将讨论金属离子对酶活性的影响。 在整个生物化学教科书,我们会经常介绍辅酶与相应的酶蛋白之间的互作。后面我们会详细 介绍维生素知识。 酶能够将一种形式的能量转化成另一种形式 很多生化反应中,反应物的能量能够高效转化成其他能量形式。例如,光合成反应能够 将光能转化成化学能。线粒体能够将食物小分子的自由能转化成离子梯度自由能,再转化成 ATP(一种通用的能量分子)。酶能够以很多方式利用 ATP 的化学键能。例如,肌球蛋白 (myosin)能够将 ATP 的能量转化成肌肉收缩的机械能(34 章)膜上的泵是一种酶,能够 利用 ATP 的能量驱动分子和离子跨膜运输(第 13 章)。化学物质和离子不对称分布所形成 的浓度梯度也是一种能量形式,有不同的用途,如发射神经脉冲。 科学家正在阐明这些能量转导酶作用的分子机制。后面章节我们将介绍不同步骤(结合、 化学转化、和释放)是如何单向循环导致能量从一种形式转化成另一种形式的机制

8.2自由能是理解酶的一个有用的热力学概念 第一章我们已经介绍了一些热力学原理。值得注意的概念是自由能(G)。为了了解酶 如何发挥作用,我们需要考虑化学反应的两个热力学性质:(1)产物和反应物之间的自由能 差异(△G):和(2)启动反应物生成产物需要的能量(即活化能△G*)。前者决定反应能否 进行,后者决定反应的速率。酶只改变后者。现在我们回顾一些能够用于酶的热力学原理。 自由能变化只提供反应能否进行的信息,不涉及反应速率 在前面我们曾经介绍,反映自由能变化△G能提供反应能否进行的信息。 1.△G小于零,反应可以发生,无需输送能量。 2.△G等于零,系统处于平衡状态。 3.△G大于零,反应不可能发生。必需输入能量驱动反应进行。因此是需能反应。 4.△G只取决于反应的终止状态(产物)和起始状态(反应物),与发生反应的路径(或 分子机理)无关。反应机理不影响反应的△G。例如氧化酶氧化葡萄糖产生CO2和H0 的自由能变化与葡萄糖直接燃烧产生CO2和H20的△G值相等。 5.△G值与反应速度没有关系。△G小于零只是表明反应能够自动发生,但是并不涉及反 应速度。反应速度取决于活化自由能(△G*)。活化自由能与△G没啥关系。 一个反应的标准自由能变化与平衡常数有关 很多反应的自由能变化可以计算,根据自由能变化确定反应是自主发生,还是需要输入 能量。为了确定这个重要的热力学参数,我们要考虑反应物和生成物的化学本质以及各个物 质的浓度。A+B==C+D △G=△G+RTn{[CD]I[AB]} (1) 其中,△G是标准自由能变化,R是气体常数,T是绝对温度,[A],B],[C],[D]是摩尔 浓度。△G是反应物和生成物浓度都是1摩尔的标准状态下自由能的变化值。因此△G取 决于反应物本质(方程式1的△G)和反应物浓度(方程式1的对数部分)。 传统上简化自由能计算。将标准状况的pH值定义为7.0。如果氢离子是反应物,其活 性(或浓度)定义为1,水的浓度(或活性)也定义为1。本书将pH7.0的自由能变化定义 为标准自由能变化(△G)。能量单位用千焦(KJ)或千卡(kcal)表示。1k=0.239kcal。 从方程1可以获得标准自由能和平衡常数之间的关系。在平衡状态,自由能变化是0, 方程1可以写成:0=△G0+RTIn{[C]D]/[AB]} (2) 那么,△G=-RTln{C]D]I[AB]} (3) 因为K'eq=[CDI/{AB]} (4) 所以△Go=-RT In K'eq (5) 即△G0=-2.303 RT log K'eq (6) 重新排列成Keq=10△G0D,303RT (7) 将R=8.315x103 kJ mol-1 deg和T=298K(相当于25℃)代入方程式7,得 K'eq=10-△G05.69 (8) 其中△G0的单位是ko1,因为必须与R的单位相匹配。因此标准自由能变化与反应的 平衡常数密切相关。例如平衡常数为10,25℃时标准自由能变化值是-5.69kJmo1(表8.3)。 注意平衡常数每变化10倍,△G0就变化5.69kJmo1
8.2 自由能是理解酶的一个有用的热力学概念 第一章我们已经介绍了一些热力学原理。值得注意的概念是自由能(G)。为了了解酶 如何发挥作用,我们需要考虑化学反应的两个热力学性质:(1)产物和反应物之间的自由能 差异(△G);和(2)启动反应物生成产物需要的能量(即活化能△G*)。前者决定反应能否 进行,后者决定反应的速率。酶只改变后者。现在我们回顾一些能够用于酶的热力学原理。 自由能变化只提供反应能否进行的信息,不涉及反应速率 在前面我们曾经介绍,反映自由能变化△G 能提供反应能否进行的信息。 1. △G 小于零,反应可以发生,无需输送能量。 2. △G 等于零,系统处于平衡状态。 3. △G 大于零,反应不可能发生。必需输入能量驱动反应进行。因此是需能反应。 4. △G 只取决于反应的终止状态(产物)和起始状态(反应物),与发生反应的路径(或 分子机理)无关。反应机理不影响反应的△G。例如氧化酶氧化葡萄糖产生 CO2和 H2O 的自由能变化与葡萄糖直接燃烧产生 CO2和 H2O 的△G 值相等。 5. △G 值与反应速度没有关系。△G 小于零只是表明反应能够自动发生,但是并不涉及反 应速度。反应速度取决于活化自由能(△G*)。活化自由能与△G 没啥关系。 一个反应的标准自由能变化与平衡常数有关 很多反应的自由能变化可以计算,根据自由能变化确定反应是自主发生,还是需要输入 能量。为了确定这个重要的热力学参数,我们要考虑反应物和生成物的化学本质以及各个物 质的浓度。 A + B ===== C + D △G = △G0 + RT ln {[C] [D] / [A] [B]} (1) 其中,△G0 是标准自由能变化,R 是气体常数,T 是绝对温度,[A],[B],[C],[D]是摩尔 浓度。△G0 是反应物和生成物浓度都是 1 摩尔的标准状态下自由能的变化值。因此△G 取 决于反应物本质(方程式 1 的△G0 )和反应物浓度(方程式 1 的对数部分)。 传统上简化自由能计算。将标准状况的 pH 值定义为 7.0。如果氢离子是反应物,其活 性(或浓度)定义为 1,水的浓度(或活性)也定义为 1。本书将 pH7.0 的自由能变化定义 为标准自由能变化(△G0 ')。能量单位用千焦(KJ)或千卡(kcal)表示。1kJ = 0.239 kcal。 从方程 1 可以获得标准自由能和平衡常数之间的关系。在平衡状态,自由能变化是 0, 方程 1 可以写成: 0 = △G0 ' + RT ln {[C] [D] / [A] [B]} (2) 那么,△G0 ' = RT ln {[C] [D] / [A] [B]} (3) 因为 K'eq = [C] [D] / {[A] [B]} (4) 所以 △G0 ' = RT ln K'eq (5) 即 △G0 ' = RT log K'eq (6) 重新排列成 K'eq = 10 △G0' /2.303 RT (7) 将 R=8.315 x 10 -3 kJ mol -1 deg -1 和 T = 298 K (相当于 25℃)代入方程式 7,得 K'eq = 10 △G0' /5.69 (8) 其中△G0 ' 的单位是 kJ mol -1,因为必须与 R 的单位相匹配。因此标准自由能变化与反应的 平衡常数密切相关。例如平衡常数为 10,25℃时标准自由能变化值是5.69 kJ mol -1(表 8.3)。 注意平衡常数每变化 10 倍,△G0 ' 就变化 5.69 kJ mol -1

TABLE 8.3 Relation between AG and K'eq (at 25C) △G Keq KJ mol- kcal mol- 10-5 28.53 6.82 104 22.84 5.46 10-3 17.11 4.09 10-2 11.42 2.73 101 5.69 1.36 1 0 0 10 -5.69 -1.36 102 -11.42 -2.73 103 -17.11 -4.09 104 -22.84 -5.46 105 -28.53 -6.82 Table 8-3 007W.H.Freemanand Compery 现在计算磷酸二羟基丙酮(DHAP)异构生成甘油醛3-磷酸(GAP)的△GO和△G。这个反 应发生在糖酵解过程中。在平衡状态,GAP与DHAP的比值是0.0475(298K,pH7.0)。 因此K'eq=0.0475。标准状态自由能变化是△G0=+7.53kJmo1 因此DHAP不能自发地生成GAP,必须输入能量才能发生。现在计算起始状态DHAP 有2x104M,GAP有3x106M的自由能变化值,△G=-2.89kmo1 结果显示DHAP能够异构生成GAP,不必输入能量。注意,尽管△GO大于零,但是 △G小于零,因此反应能够自发进行。无论△G比△G大、小、还是相等,反应能否自动 进行取决于△G,而不是△G。这,点必须记住。那些根据△G不能自发进行的反映可以 通过调节反应物和生成物的浓度使之能够发生。这一原则是偶联化学反应形成代谢途径的基 础(第15章)。 酶只改变化学反应速度,并不改变化学反应平衡 酶是超级催化剂,诱使人们觉得酶具有它们自身并不具有的能力。实际上,酶不能改变 热力学法则,因此不能改变反应平衡。考察一个酶将底物S转化成产物P,用产物形成量对 时间作图(图82)。有酶,化学反应几秒钟就产生了没有酶时需要几小时甚至几年才能形成 的产量。但是我们注意到产物的最终产量不受酶影响。 Enzyme onpold No enzyme Seconds Hours Time 品 图82酶加速化学反应。无论有没有酶,化学反应都能达到同一平衡点。但是酶能够加速反 应,迅速达到反应平衡
现在计算磷酸二羟基丙酮(DHAP)异构生成甘油醛 3-磷酸(GAP)的△G0 ' 和△G。这个反 应发生在糖酵解过程中。在平衡状态,GAP 与 DHAP 的比值是 0.0475 (298K, pH7.0)。 因此 K'eq = 0.0475。标准状态自由能变化是△G0 ' = + 7.53 kJ mol -1 因此 DHAP 不能自发地生成 GAP,必须输入能量才能发生。现在计算起始状态 DHAP 有 2 x 10 -4 M, GAP 有 3 x 10 -6 M 的自由能变化值,△G = - 2.89 kJ mo l-1 结果显示 DHAP 能够异构生成 GAP,不必输入能量。注意,尽管△G0 ' 大于零,但是 △G 小于零,因此反应能够自发进行。无论△G 比△G0 ' 大、小、还是相等,反应能否自动 进行取决于△G,而不是△G0 ' 。这点必须记住。那些根据△G0 ' 不能自发进行的反映可以 通过调节反应物和生成物的浓度使之能够发生。这一原则是偶联化学反应形成代谢途径的基 础(第 15 章)。 酶只改变化学反应速度,并不改变化学反应平衡 酶是超级催化剂,诱使人们觉得酶具有它们自身并不具有的能力。实际上,酶不能改变 热力学法则,因此不能改变反应平衡。考察一个酶将底物 S 转化成产物 P,用产物形成量对 时间作图(图 8.2)。有酶,化学反应几秒钟就产生了没有酶时需要几小时甚至几年才能形成 的产量。但是我们注意到产物的最终产量不受酶影响。 图 8.2 酶加速化学反应。无论有没有酶,化学反应都能达到同一平衡点。但是酶能够加速反 应,迅速达到反应平衡