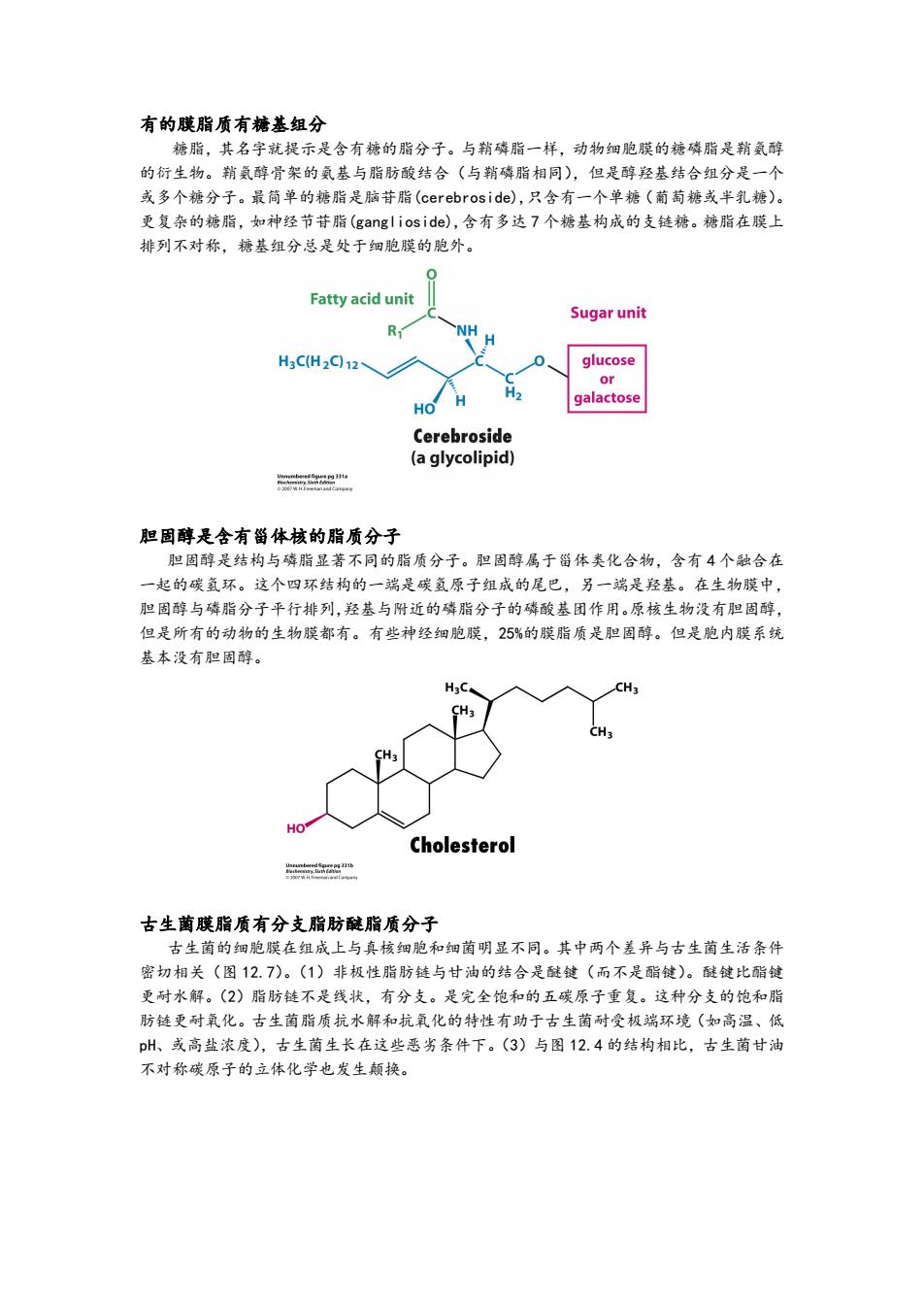
有的膜脂质有糖基组分 糖脂,其名字就提示是含有糖的脂分子。与鞘磷脂一样,动物细胞膜的糖磷脂是鞘氨醇 的衍生物。鞘氨醇骨架的氨基与脂肪酸结合(与鞘磷脂相同),但是醇羟基结合组分是一个 或多个糖分子。最简单的糖脂是脑苷脂(cerebros ide),只含有一个单糖(蔺萄糖或半乳糖)。 更复杂的糖脂,如神经节苷脂(ganglioside),含有多达7个糖基构成的支链糖。糖脂在膜上 排列不对称,糖基组分总是处于细胞膜的胞外。 0 Fatty acid unit Sugar unit R NH H3CH2q12、 glucose or galactose HO Cerebroside (a glycolipid) 胆固醇是含有甾体核的脂质分子 胆固醇是结构与磷脂显著不同的脂质分子。胆固醇属于甾体类化合物,含有4个融合在 一起的碳氢环。这个四环结构的一端是碳氢原子组成的尾巴,另一端是羟基。在生物膜中, 胆固醇与磷脂分子平行排列,羟基与附近的磷脂分子的磷酸基团作用。原核生物没有胆固醇, 但是所有的动物的生物膜都有。有些神经细胞膜,25%的膜脂质是胆固醇。但是胞内膜系统 基本没有胆固醇。 H3C CH3 CH3 CH3 HO Cholesterol 古生菌膜脂质有分支脂肪醚脂质分子 古生菌的细胞膜在组成上与真核细胞和细菌明显不同。其中两个差异与古生菌生活条件 密切相关(图12.7)。(1)非极性脂肪链与甘油的结合是醚键(而不是酯键)。醚键比酯键 更耐水解。(2)脂肪链不是线状,有分支。是完全饱和的五碳原子重复。这种分支的饱和脂 肪链更耐氧化。古生菌脂质抗水解和抗氧化的特性有助于古生菌耐受极端环境(如高温、低 pH、或高盐浓度),古生菌生长在这些恶劣条件下。(3)与图12.4的结构相比,古生菌甘油 不对称碳原子的立体化学也发生颠换
有的膜脂质有糖基组分 糖脂,其名字就提示是含有糖的脂分子。与鞘磷脂一样,动物细胞膜的糖磷脂是鞘氨醇 的衍生物。鞘氨醇骨架的氨基与脂肪酸结合(与鞘磷脂相同),但是醇羟基结合组分是一个 或多个糖分子。最简单的糖脂是脑苷脂(cerebroside),只含有一个单糖(葡萄糖或半乳糖)。 更复杂的糖脂,如神经节苷脂(ganglioside),含有多达 7 个糖基构成的支链糖。糖脂在膜上 排列不对称,糖基组分总是处于细胞膜的胞外。 胆固醇是含有甾体核的脂质分子 胆固醇是结构与磷脂显著不同的脂质分子。胆固醇属于甾体类化合物,含有 4 个融合在 一起的碳氢环。这个四环结构的一端是碳氢原子组成的尾巴,另一端是羟基。在生物膜中, 胆固醇与磷脂分子平行排列,羟基与附近的磷脂分子的磷酸基团作用。原核生物没有胆固醇, 但是所有的动物的生物膜都有。有些神经细胞膜,25%的膜脂质是胆固醇。但是胞内膜系统 基本没有胆固醇。 古生菌膜脂质有分支脂肪醚脂质分子 古生菌的细胞膜在组成上与真核细胞和细菌明显不同。其中两个差异与古生菌生活条件 密切相关(图 12.7)。(1)非极性脂肪链与甘油的结合是醚键(而不是酯键)。醚键比酯键 更耐水解。(2)脂肪链不是线状,有分支。是完全饱和的五碳原子重复。这种分支的饱和脂 肪链更耐氧化。古生菌脂质抗水解和抗氧化的特性有助于古生菌耐受极端环境(如高温、低 pH、或高盐浓度),古生菌生长在这些恶劣条件下。(3)与图 12.4 的结构相比,古生菌甘油 不对称碳原子的立体化学也发生颠换

图12.7一种古生菌及其生活环境。古生菌可以生活在火山口这样的极端环境中。此处的古 生菌形成橘红色垫子,周围是黄色含硫污泥。 0 H.C H Membrane lipid from the archaeon Methanococcus jannaschii 高 膜脂质是含有一个亲水区和一个疏水区的两性分子 膜脂质分子式样很多,但是它们有共同的结构特征。膜脂质是两性分子(amphilic molecule)。生物膜的一个脂质分子既含有亲水区域也含有疏水区域。 现在来看看磷脂酰甘油酯(如磷脂酰胆碱)的结构模型。这种脂质分子的总结构形状大 致是长方形(图12.8A)。两个疏水性脂肪酸链相互间大致平行排列,而亲水性磷酰胆碱区 域指向另一侧。鞘磷脂的构型与此类似,古生菌的脂质分子也是这样。因此,脂质分子可以 简写为,圆圈代表极性的亲水头部,直线或波浪线代表疏水的碳氢尾巴(图12.8B)。 Phosphoglyceride ●●●◆● Sphingomyelin Archaeal lipid Shorthand depiction 图12.8膜脂质示意图。(A)磷脂酰甘油脂、鞘磷脂、和古生菌脂质分子的空间填充模型, 显示这些脂质分子的形状,亲水部分和疏水部分在分子结构上的分布。(B)膜脂质分子的结 构缩写
图 12.7 一种古生菌及其生活环境。古生菌可以生活在火山口这样的极端环境中。此处的古 生菌形成橘红色垫子,周围是黄色含硫污泥。 膜脂质是含有一个亲水区和一个疏水区的两性分子 膜脂质分子式样很多,但是它们有共同的结构特征。膜脂质是两性分子(amphilic molecule)。生物膜的一个脂质分子既含有亲水区域也含有疏水区域。 现在来看看磷脂酰甘油酯(如磷脂酰胆碱)的结构模型。这种脂质分子的总结构形状大 致是长方形(图 12.8A)。两个疏水性脂肪酸链相互间大致平行排列,而亲水性磷酰胆碱区 域指向另一侧。鞘磷脂的构型与此类似,古生菌的脂质分子也是这样。因此,脂质分子可以 简写为,圆圈代表极性的亲水头部,直线或波浪线代表疏水的碳氢尾巴(图 12.8B)。 图 12.8 膜脂质示意图。(A)磷脂酰甘油脂、鞘磷脂、和古生菌脂质分子的空间填充模型, 显示这些脂质分子的形状,亲水部分和疏水部分在分子结构上的分布。(B)膜脂质分子的结 构缩写

12.3磷脂和糖脂在水相能形成双分子片层结构 磷脂的什么性质能形成膜?脂质分子的两性特征导致膜的形成。在水溶液中脂质分子的 极性头部喜好与水接触,而非极性尾部相互接触。有这种倾向性的分子在水溶液中如何排 列?一种结构是形成微团(micel le)。极性头部处于微团表面,周围是水分子,疏水性的烃 类尾巴相互接触、包埋于微团内部(图12.9)。 图12.9微团的截面图谱。解离的脂肪酸易于形成这样的结构,但是大多数磷脂分子不形成 这样的结构。 膜脂质分子完全相反的溶解选择性倾向于形成两层脂质分子构成的脂质双层(图 12.10)。脂质双层也叫双分子层。一层脂质分子的疏水性尾部与另一层脂质分子的疏水性尾 部接触,形成疏水性内部,阻碍分子透过。双分子层各面的极性头部与水相接触。每个脂质 双层的两个对应的分子层称为小叶(leaflet)。 图12.10双层膜的截面示意图。 在水相环境中,大多数磷脂或糖脂所形成的结构是双分子脂质层,而不是微团。原因是 磷脂或糖脂的两个脂肪酸链太大,微团内部空间容纳不下。相反,脂肪酸(如软脂酸钠,肥 皂的组分)易于形成微泡,因为这些分子只有一条脂肪链。磷脂分子形成脂质双层(而不形 成微泡)有重要的生物学意义。微泡大小有限,通常直径不超过20m。相反,直至双层的 片状结构能够扩展球体的直径,直径可达1毫米(10°m)甚至更大。磷脂和相关的生物分子 是生物膜重要组分,因为这些分子易于形成片状的脂质双层结构(图12.11)。 分子自组装能够形成脂质双层。换句话说,含有脂质分子都能形成脂质双层结构。在水 中,磷脂分子自动形成脂质双层的速度快。疏水作用是形成脂质双层的主要驱动力。我们记 得,疏水作用在核酸碱基堆积和蛋白质折叠过程中作用显著。膜脂质分子疏水性尾巴置于脂 质双层内部的非极性环境,释放水分子。而且疏水尾巴的van der Waals相互作用有助于它
12.3 磷脂和糖脂在水相能形成双分子片层结构 磷脂的什么性质能形成膜?脂质分子的两性特征导致膜的形成。在水溶液中脂质分子的 极性头部喜好与水接触,而非极性尾部相互接触。有这种倾向性的分子在水溶液中如何排 列?一种结构是形成微团(micelle)。极性头部处于微团表面,周围是水分子,疏水性的烃 类尾巴相互接触、包埋于微团内部(图 12.9)。 图 12.9 微团的截面图谱。解离的脂肪酸易于形成这样的结构,但是大多数磷脂分子不形成 这样的结构。 膜脂质分子完全相反的溶解选择性倾向于形成两层脂质分子构成的脂质双层(图 12.10)。脂质双层也叫双分子层。一层脂质分子的疏水性尾部与另一层脂质分子的疏水性尾 部接触,形成疏水性内部,阻碍分子透过。双分子层各面的极性头部与水相接触。每个脂质 双层的两个对应的分子层称为小叶(leaflet)。 图 12.10 双层膜的截面示意图。 在水相环境中,大多数磷脂或糖脂所形成的结构是双分子脂质层,而不是微团。原因是 磷脂或糖脂的两个脂肪酸链太大,微团内部空间容纳不下。相反,脂肪酸(如软脂酸钠,肥 皂的组分)易于形成微泡,因为这些分子只有一条脂肪链。磷脂分子形成脂质双层(而不形 成微泡)有重要的生物学意义。微泡大小有限,通常直径不超过 20 nm。相反,直至双层的 片状结构能够扩展球体的直径,直径可达 1 毫米(10 6 nm)甚至更大。磷脂和相关的生物分子 是生物膜重要组分,因为这些分子易于形成片状的脂质双层结构(图 12.11)。 分子自组装能够形成脂质双层。换句话说,含有脂质分子都能形成脂质双层结构。在水 中,磷脂分子自动形成脂质双层的速度快。疏水作用是形成脂质双层的主要驱动力。我们记 得,疏水作用在核酸碱基堆积和蛋白质折叠过程中作用显著。膜脂质分子疏水性尾巴置于脂 质双层内部的非极性环境,释放水分子。而且疏水尾巴的 van der Waals 相互作用有助于它
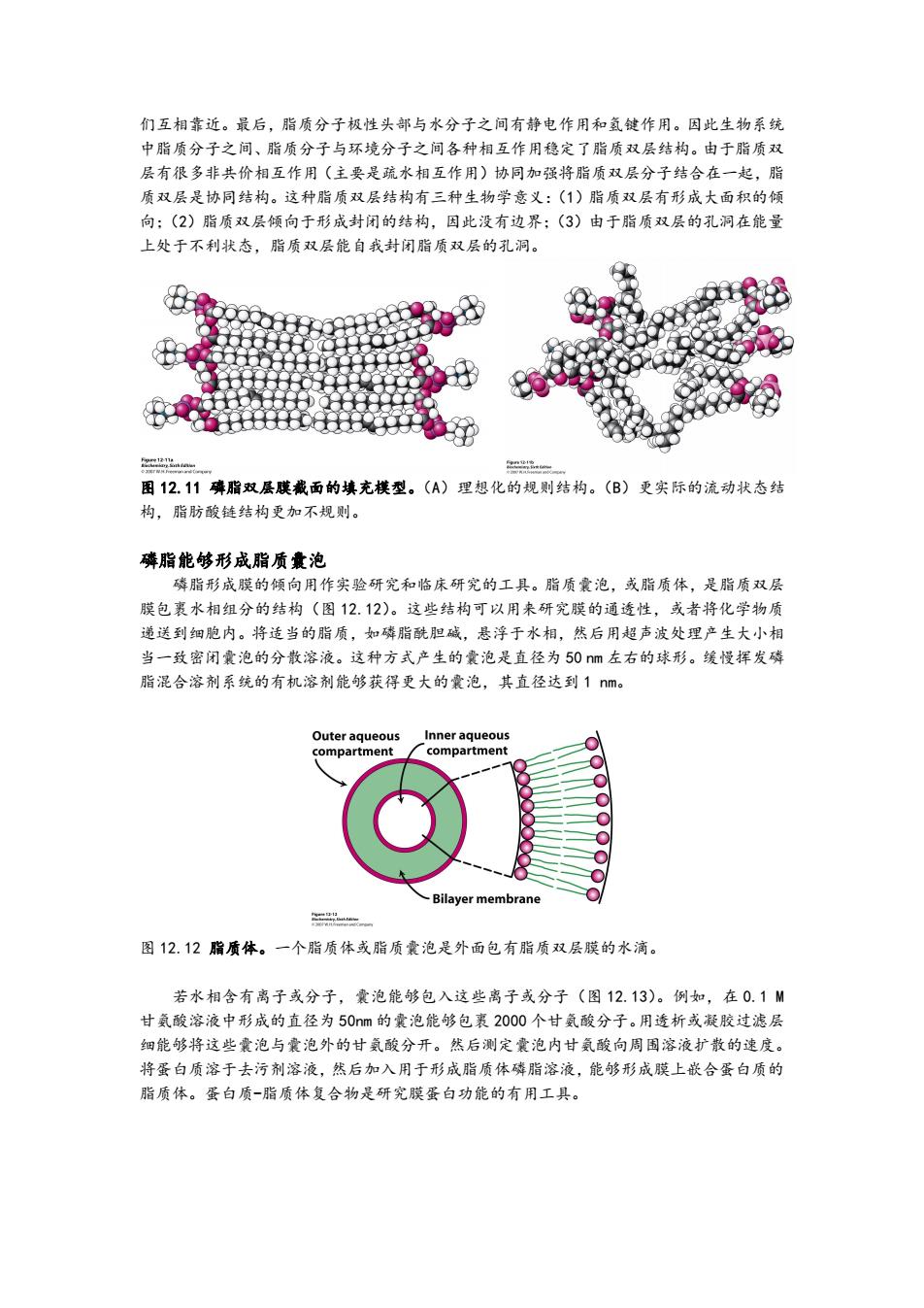
们互相靠近。最后,脂质分子极性头部与水分子之间有静电作用和氢键作用。因此生物系统 中脂质分子之间、脂质分子与环境分子之间各种相互作用稳定了脂质双层结构。由于脂质双 层有很多非共价相互作用(主要是疏水相互作用)协同加强将脂质双层分子结合在一起,脂 质双层是协同结构。这种脂质双层结构有三种生物学意义:(1)脂质双层有形成大面积的倾 向:(2)脂质双层倾向于形成封闭的结构,因此没有边界:(3)由于脂质双层的孔洞在能量 上处于不利状态,脂质双层能自我封闭脂质双层的孔洞。 2Ce 图12.11磷脂双层膜戴面的填充模型。()理想化的规则结构。(B)更实际的流动状态结 构,脂肪酸链结构更加不规则。 璘脂能够形成脂质囊泡 磷脂形成膜的倾向用作实验研究和临床研究的工具。脂质囊泡,或脂质体,是脂质双层 膜包裹水相组分的结构(图12.12)。这些结构可以用来研究膜的通透性,或者将化学物质 递送到细胞内。将适当的脂质,如磷脂酰胆碱,悬浮于水相,然后用超声波处理产生大小相 当一致密闭囊泡的分散溶液。这种方式产生的囊泡是直径为50m左右的球形。缓慢挥发磷 脂混合溶剂系统的有机溶剂能够获得更大的囊泡,其直径达到1nm。 Outer aqueous Inner aqueous compartment compartment -Bilayer membrane 二 图12.12脂质体。一个脂质体或脂质囊泡是外面包有脂质双层膜的水滴。 若水相含有离子或分子,囊泡能够包入这些离子或分子(图12.13)。例如,在0.1M 甘氨酸溶液中形成的直径为50m的囊泡能够包裹2000个甘氨酸分子。用透析或凝胶过滤层 细能够将这些囊泡与囊泡外的甘氨酸分开。然后测定囊泡内甘氨酸向周围溶液扩散的速度。 将蛋白质溶于去污剂溶液,然后加入用于形成脂质体磷脂溶液,能够形成膜上嵌合蛋白质的 脂质体。蛋白质一脂质体复合物是研究膜蛋白功能的有用工具
们互相靠近。最后,脂质分子极性头部与水分子之间有静电作用和氢键作用。因此生物系统 中脂质分子之间、脂质分子与环境分子之间各种相互作用稳定了脂质双层结构。由于脂质双 层有很多非共价相互作用(主要是疏水相互作用)协同加强将脂质双层分子结合在一起,脂 质双层是协同结构。这种脂质双层结构有三种生物学意义:(1)脂质双层有形成大面积的倾 向;(2)脂质双层倾向于形成封闭的结构,因此没有边界;(3)由于脂质双层的孔洞在能量 上处于不利状态,脂质双层能自我封闭脂质双层的孔洞。 图 12.11 磷脂双层膜截面的填充模型。(A)理想化的规则结构。(B)更实际的流动状态结 构,脂肪酸链结构更加不规则。 磷脂能够形成脂质囊泡 磷脂形成膜的倾向用作实验研究和临床研究的工具。脂质囊泡,或脂质体,是脂质双层 膜包裹水相组分的结构(图 12.12)。这些结构可以用来研究膜的通透性,或者将化学物质 递送到细胞内。将适当的脂质,如磷脂酰胆碱,悬浮于水相,然后用超声波处理产生大小相 当一致密闭囊泡的分散溶液。这种方式产生的囊泡是直径为 50 nm 左右的球形。缓慢挥发磷 脂混合溶剂系统的有机溶剂能够获得更大的囊泡,其直径达到 1 nm。 图 12.12 脂质体。一个脂质体或脂质囊泡是外面包有脂质双层膜的水滴。 若水相含有离子或分子,囊泡能够包入这些离子或分子(图 12.13)。例如,在 0.1 M 甘氨酸溶液中形成的直径为 50nm 的囊泡能够包裹 2000 个甘氨酸分子。用透析或凝胶过滤层 细能够将这些囊泡与囊泡外的甘氨酸分开。然后测定囊泡内甘氨酸向周围溶液扩散的速度。 将蛋白质溶于去污剂溶液,然后加入用于形成脂质体磷脂溶液,能够形成膜上嵌合蛋白质的 脂质体。蛋白质-脂质体复合物是研究膜蛋白功能的有用工具