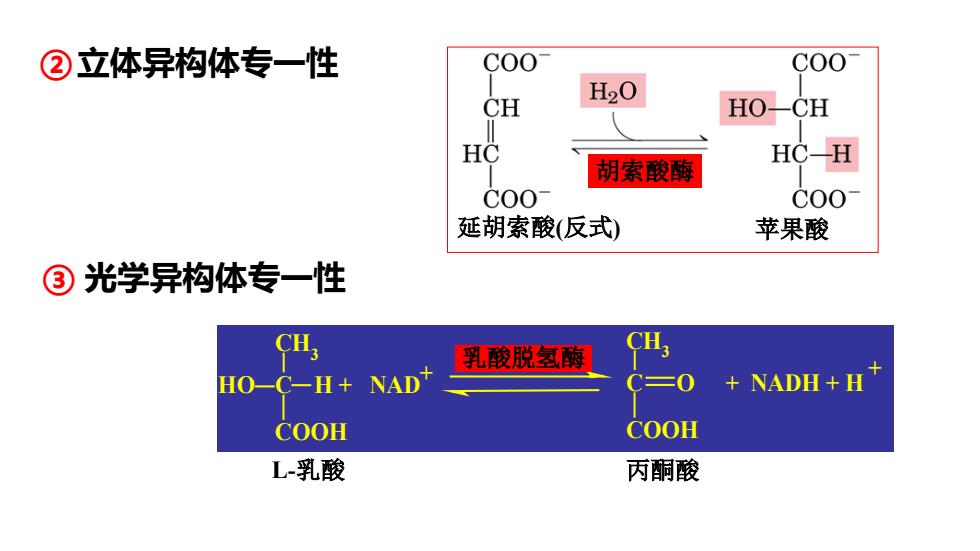
②立体异构体专一性 C00 CH H20 HO-CH HC HC-H 胡索酸酶 C00 C00 延胡索酸(反式) 苹果酸 ③光学异构体专一性 CH 乳酸脱氢酶 CH. HO- C一H+ NAD 三0 NADH+H COOH COOH L-乳酸 丙酮酸
②立体异构体专一性 胡索酸酶 延胡索酸(反式) 苹果酸 CH3 C COOH O CH3 C COOH HO H + NAD + NADH + H + LDH + L-乳酸 丙酮酸 乳酸脱氢酶 ③ 光学异构体专一性

3.可调节性: √酶促反应受多种因素的调控,以适应机体对不断变化的内外环 境和生命活动的需要。 √酶的调节方式包括酶含量和酶活性的调节。 √基本模式:各种细胞内外因素→调节酶的活性和含量→调节 代谢等过程。 4.不稳定性: √酶是蛋白质等生物大分子,故其作用条件比较温和(中性H、 常温和常压),强酸、强碱、高温条件均易使酶发生变性而失 去活性
3. 可调节性: 酶促反应受多种因素的调控,以适应机体对不断变化的内外环 境和生命活动的需要。 酶的调节方式包括酶含量和酶活性的调节。 基本模式:各种细胞内外因素 → 调节酶的活性和含量 → 调节 代谢等过程。 4.不稳定性: 酶是蛋白质等生物大分子,故其作用条件比较温和(中性pH、 常温和常压),强酸、强碱、高温条件均易使酶发生变性而失 去活性

二、酶促反应的机制 (工作原理) 1.酶比一般催化剂能更有效地降低反应活化能。 2.诱导契合假说(1958年,Daniel Koshland等提出) : √是指酶与底物接近时,其结构相互诱导、相互变形和相互适应, 进而相互结合,形成E-S复合物。 √不是机械的锁钥模式 ,(1894年Emil Fischer提出)。 底物 酶活中心 化学键 辅助因子 游离酶 过菠态复合物 B D 底物结合位点 酶底复合物 原始酶
二、酶促反应的机制(工作原理) 1. 酶比一般催化剂能更有效地降低反应活化能。 2. 诱导契合假说(1958年,Daniel Koshland等提出): 是指酶与底物接近时,其结构相互诱导、相互变形和相互适应, 进而相互结合,形成E-S复合物。 不是机械的锁-钥模式(1894年Emil Fischer提出)
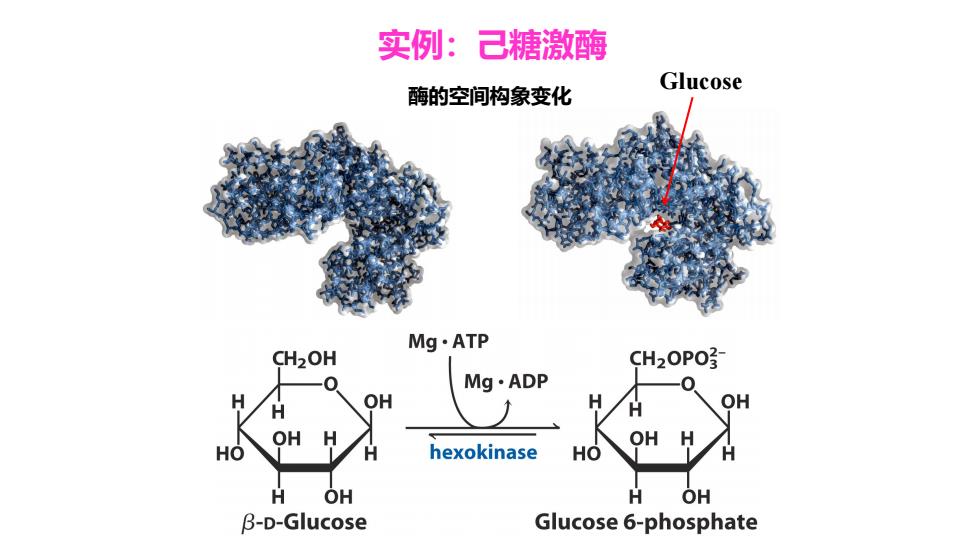
实例:己糖激酶 酶的空间构象变化 Glucose Mg·ATP CH2OH CH2OPO? Mg·ADP OH H OH OH HO hexokinase HO OH B-D-Glucose Glucose 6-phosphate
实例:己糖激酶 酶的空间构象变化 Glucose

3.酶催化机制的多元性(了解): √一种酶催化的反应往往是多种催化机制的综合作用,这是酶促 反应高效率的重要原因。 √邻近效应与定向效应:增加酶与底物的接触机会和有效碰撞 √多元催化:酶是两性电解质,含多种功能基团 √表面效应:酶活性中兴多为内陷疏水口袋 √张力和变形作用:诱导底物变形扭曲,促进键断裂 / 共价催化:酶与底物形成过渡性的共价中间体,限制底物的活动,使反 应易于进行
3. 酶催化机制的多元性(了解): 一种酶催化的反应往往是多种催化机制的综合作用,这是酶促 反应高效率的重要原因。 邻近效应与定向效应:增加酶与底物的接触机会和有效碰撞 多元催化:酶是两性电解质,含多种功能基团 表面效应:酶活性中兴多为内陷疏水口袋 张力和变形作用:诱导底物变形扭曲,促进键断裂 共价催化:酶与底物形成过渡性的共价中间体,限制底物的活动,使反 应易于进行