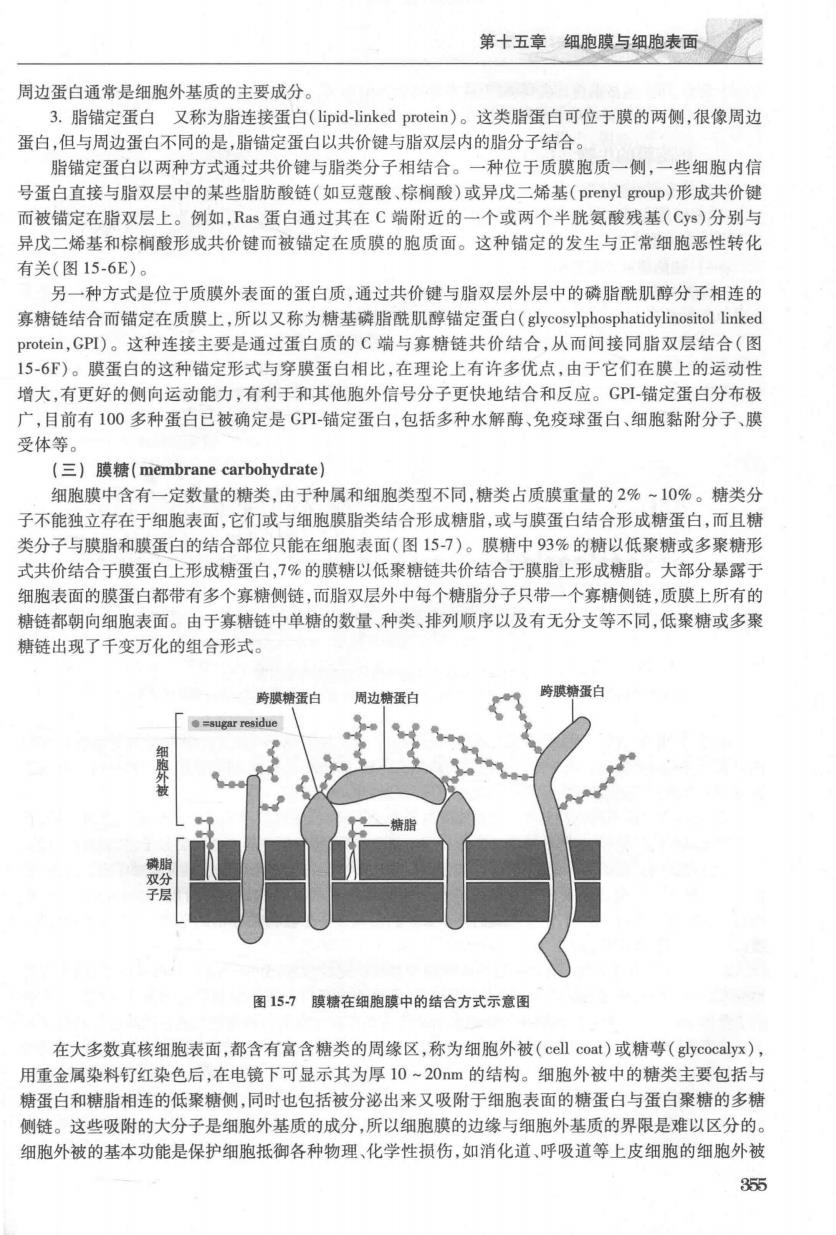
第十五章细胞膜与细胞表面 周边蛋白通常是细胞外基质的主要成分。 3.脂锚定蛋白又称为脂连接蛋白(lipid-linked protein)。这类脂蛋白可位于膜的两侧,很像周边 蛋白,但与周边蛋白不同的是,脂错定蛋白以共价键与脂双层内的脂分子结合 脂锚定蛋白以两种方式通过共价键与脂类分子相结合。 一种位于质膜胞质一侧,一些细胞内信 号蛋白直接与脂双层中的某些脂肪酸链(如豆蔻酸、棕榈酸)或异戊二烯基(prenyl group)形成共价键 而被铺定在脂双层上。例如,Ras蛋白通过其在C端附近的一个或两个半胱氨酸残基(Cy)分别与 异戊二烯基和棕榈酸形成共价键而被错定在质膜的胞质面。这种错定的发生与正常细胞恶性转化 有关(图15-6E) 另一种方式是位于质莫外表面的蛋白质,通过共价键与脂双层外层中的磷脂酰肌醇分子相连的 毫糖链结合而错定在质膜上,所以又称为糖基陵脂酰肌醇错定蛋白(glvcosylphosphatidvlinositol linked protein,GP)。这种连接主要是通过蛋白质的C端与寡糖链共价结合,从而间接同脂双层结合(图 15-6F)。膜蛋白的这种锚定形式与穿膜蛋白相比,在理论上有许多优点,由于它们在膜上的运动性 增大,有更好的侧向运动能力,有利于和其他胞外信号分子更快地结合和反应。G-错定蛋白分布极 广,目前有100多种蛋白已被确定是GPI错定蛋白,包括多种水解酶、免疫球蛋白、细胞黏附分子、膜 受体等 (三)膜糖(membrane carbohydrate】 细胞膜中含有一定数量的糖类,由于种属和细胞类型不同,糖类占质膜重量的2%~10%。糖类分 子不能独立存在于细胞表面,它们或与细胞膜脂类结合形成糖脂,或与膜蛋白结合形成糖蛋白,而且糖 类分子与膜脂和膜蛋白的结合部位只能在细胞表面(图15-7)。膜糖中93%的糖以低聚糖或多聚糖形 式共价结合干膜蛋白上形成糖蛋白,7%的膜糖以低聚糖链共价结合于膜脂上形成糖脂。大部分暴碳于 细胞表面的膜蛋白都带有多个寡糖侧链,而脂双层外中每个糖脂分子只带一个寡糖侧链,质膜上所有的 糖链都朝向细胞表面。由于寡糖链中单糖的数量、种类、排列顺序以及有无分支等不同,低聚糖或多聚 糖链出现了千变万化的组合形式。 周边糖蛋白 跨膜精蛋 图15-7膜精在细胞膜中的结合方式示意国 在大多数真核细胞表面,都含有富含糖类的周缘区,称为细胞外被(cel1coat)或糖尊(yeocalyx), 用重金属染料钉红染色后,在电镜下可显示其为厚10~20m的结构。细胞外被中的糖类主要包括与 糖蛋白和糖脂相连的低聚糖侧,同时也包括被分泌出来又吸附于细胞表面的糖蛋白与蛋白聚糖的多树 侧链。这些吸附的大分子是细胞外基质的成分,所以细胞膜的边缘与细胞外基质的界限是难以区分的。 细胞外被的基本功能是保护细胞抵御各种物理,化学性损伤,如消化道,呼吸道等上皮细胞的细胞外被 355

第四篇细胞的结构与功能 有助于润滑、防止机械损伤、保护黏膜上皮不受消化酶的作用。糖链末端富含带负电荷的唾液酸,能捕 集Na、C'等阳离子并吸引大量的水分子,使细胞周围建立起水盐平衡的微环境。 二、细胞膜的生物学特性 细胞膜由脂双分子层和以不同方式与其结合的蛋白质组成,它不仅具有保护细胞的“屏障”的作 用,还具有物质运输、信号传递、细胞识别等多种重要功能,细胞膜的这一系列生物学功能都与细胞膜的 分子结构和组成特性有关。细胞膜具有两个显著的生物学特性,即膜的不对称性和流动性。 (一)细胞膜的不对称性 细胞膜内外两层在结构和功能上具有很大差异性,称为膜的不对称性(membrane asymmetry),包括细 胞膜中各种组成成分的不对称和功能的不对称,细胞膜组成的不对称决定了其功能的不对称性和方向性。 1,膜脂的不对称性实验检测各种膜脂双层的化学组成,发现各种膜脂在脂双层内、外两单层的 分布是不相同的,同一种膜脂分子在脂双层中呈不均匀分布。例如,在人红细胞膜中,绝大部分的鞘磷 脂、卵磷脂和胆固醇多位于脂双层外层,而磷脂酰乙醇胺、磷脂酰丝氨酸和磷脂酰肌醇在内层中含量较 多,并且不同的膜脂在含量和比例上也存在较大差异(图15-8)。 细胞外 细胞内 图158膜脂在细胞膜中不对称分布示意图 SM:鞘磷脂:P刊:磷脂酰肌醇:CI:胆固醇:PS:磷脂酰丝氨酸:PC:卵磷脂:PE:磷脂酰乙醇胺 此外,不同膜性细胞器中脂类成分的组成和分布不同。如质膜中一般富含鞘磷脂、磷脂酰胆碱和 固醇等:核膜、内质网膜和线粒体外膜则富含磷脂酰胆碱、磷脂酰乙醇胺、磷脂酰肌醇。正是由于存在膜 脂成分分布的差异,使细胞内的生物膜具有不同的特性和功能。 目前,脂质双层不对称性分布的生理意义尚不完全明确,但它可以使膜的两层流动性有所不同,有 助于维持膜蛋白的极性,与膜的特定功能相一致。研究发现,脂双层中的一些脂类分子,如磷脂酰肌醇 可以为特定蛋白质提供结合位点,对保持膜蛋白在脂双层中的正确定位和极性具有重要作用。此外,膜 脂的不对称性还表现在膜表面具有胆固醇和鞘磷脂等形成的微结构域一脂筏(lipid raft),大小约 70m左右,是一种动态结构,位于质膜的外小页。脂筏就像一个蛋白质停泊的平台,与膜的信号转导 蛋白质分选均有密切的关系。 2.膜蛋白的不对称性各种膜蛋白在质膜中都有一定的位置,分布于脂双层内外两层,属于不对 称分布。不同蛋白质在细胞膜上的位置是不同的,例如血影蛋白分布于细胞膜内侧面;磷酸酯酶、生长 因子受体、激素受体等位于质膜的外侧面:腺苷酸环化酶则位于质膜的内侧胞质面。用冰冻蚀刻技术显 示细胞膜的两个剖面,可以清楚地看到膜蛋白在脂双层内、外两层中的分布有明显差异。此外,跨膜蛋 白穿越脂双层都有一定的方向性,这也造成其分布的不对称性。例如,红细胞膜上的血型糖蛋白肽链的 N端伸向质膜外侧,C端在质膜内侧胞质面;而带Ⅲ蛋白肽链的N端则在质膜内侧胞质面。膜蛋白的不 对称性还表现在跨膜蛋白的两个亲水端,其肽链长度、氨基酸的种类和顺序都不同,有的在膜外侧有作 用位点,有的在膜内侧有作用位点。糖蛋白的糖残基均分布在生物膜的非胞质一侧(图159)。 356

第十五章细胞膜与细胞表面人 3.膜糖的不对称性细胞膜所含的糖类 只能与膜脂或膜蛋白结合,以糖脂和糖蛋白的 形式存在于细胞膜上,因此,膜糖类的分布具 i0iimM0i0iN 有显著的不对称性。细胞膜糖脂、糖蛋白的寡 糖侧鞋只分布于质膜外表面,而在胞质中各膜 性细胞器的寡糖侧链都分布于膜腔的内侧面。 膜脂、膜蛋白及膜糖分布的不对称性与膜功能 跨膜蛋 周边蛋白 的不对称性和方向性有密切关系,具有重要的 生物学意义,膜结构上的不对称性保证了膜功 图159膜蛋白在细胞膜中分布的不对称性 能的方向性和生命活动的高度有序性。例如 S:细胞外表面;EF:细胞外小页断裂面;PS:细胞 人红细胞膜表面的寒糖特决定了人举的AB0 内表面:P℉:细胞内小页断面 血型;研究发现多数激素的受体都位于质膜的 外侧,接受细胞外信号并向细胞内传递:当细胞发生调亡时,原本位于脂双层内侧的磷脂酰丝氨酸会翻 转到外层,成为巨噬细胞识别并吞噬调亡细胞的信号。 (二)细胞膜的流动性 膜的流动性(iiy)保证细胞膜进行正常的功能活动,是细胞进行生命活动的必需条件,也是细胞 膜的基本特性之一。细胞膜是一个动态结构,其流动性包括膜脂的流动性和膜蛋白的流动性 1.膜脂的流动性研究认为组成细胞膜基本骨架的脂双层既有固体分子排列的有序性,又有液体 的流动性,这一兼有晶态和液态两种特性的状态称为液晶态(liquid-crystal state),是细胞膜极为重要的 特性。在生理条件下,膜大多处于液晶态。在温度下降到一定程度,脂双层的性质会明显改变,从流动 的液晶态转变为“冰冻”的晶品态(或凝胶态),这是磷脂分子的运动将受到很大限制:当温度上升到某 点时,晶态也可以熔融为液晶态,把引起相变的这一临界温度称为膜的相变温度。由于温度的变化导致 膜状态的改变称为相变(phase transition)。在相变温度以上,膜处于流动的液晶态。 研究表明,脂类分子的各种运动与膜结构的流动性密切相关,在高于相变温度的条件下,膜脂分子 的运动方式可以归纳为以下几种(图15-10)。 侧向扩散旋转运动 弯曲运动 转运动 旋转异构 图15-10膜脂分子在细胞膜中几种运动方式 (I)侧向扩散(lateral diffusion):侧向扩散是指在脂双层的同一单分子层内,脂分子沿膜平面侧向 与相邻分子快速交换位置,其交换频率在10次/s以上。这种快速横向扩散运动以扩散系数表示, 一般 定义为处于液晶态的脂双层在30℃时其脂类分子的则向扩散系数(D)10m/s,即每秒移动2um的距 离。侧向扩散运动是膜脂分子主要的运动方式,具有重要的生物学意义。侧向扩散始终保持脂类分子 的排列方向,亲水的头部基团朝向膜表面,疏水的尾部朝向膜的内部。 (2)旋转运动(rotation):是膜脂分子围绕与膜平面相垂直的轴进行自旋运动,约10~10/s。 (3)弯曲运动(1exio):膜脂分子的烃链是有韧性和可弯曲的,分子的尾部端弯曲、摆动幅度大,而 靠近极性头部的弯曲摆动幅度小

第四篇细胞的结构与功能 (4)伸缩震荡(stretching shock):是脂肪酸链沿着与膜平面垂直的长轴进行伸缩、振荡运动。 (5)翻转运动(-p):是膜脂分子在双分子层之间由脂双层的一层翻转至另一层的运动。这种 运动一般情况下很少发生,因为当发生翻转运动时,麟脂的亲水头部基团将穿过膜内部的硫水层,克服 硫水区的阻力方能抵达另一个层面,这在热力学上是很不利的。但是脂质分子的翻转运动在内膜系统 内质网上发生频率很高,内质网膜上有一种翻转酶(lippase),它能促使某些新合成的磷脂分子从脂双 层的胞质面翻转到非胞质面。这些酶在维持膜脂的不对称分布中起重要作用。 (6)旋转异构运动:指烃链C-C健的旋转引起的异构运动,使烃链可以在全反式构象和歪扭构 之间产生旋转。在低温条件下,烃链呈全反式构象,随着温度升高,歪扭构象逐渐增多,流动性加大。这 种旋转异构运动的速度很快,其跳动頫率为10~10°/s 膜的流动型是膜功能活动的重要保证。有了膜的流动性,膜蛋白可以在膜的特定位点聚集形成特 定结构或功能单位,以完成如细胞连接建立、信号转导等多种功能活动。许多基本的生命活动,包括细 胞的运动、生长分裂、物质转运、分泌和吞噬等作用,都取决于膜组分的运动。如果膜是一种刚性、非沙 态结构,这些行为都不可能发生。 2.影响膜脂流动性的因素膜脂的流动性对于膜的功能具有重要作用,脂双层的流动性主要依赖 于其组分和脂分子本身的结构特征,影响其流动性的主要因素如下(图15-11): 流动性增加 图1511影响膜脂流动性的因素(示脂肪酸链的饱和程度与胆固醇 A.脂肪酸链的饱和程度与膜脂流动性:B.胆固醇分子与膜脂流动性 (1)脂肪酸链的饱和程度:研究发现相变温度的高低和流动性的大小决定于脂类分子排列的紧密 程度。避脂分子长的询和脂肪酸靠县吉线形,且右最大的廉集倾向而排列成紧密的疑胶状态:而不饱利 脂肪酸链在双健处形成折屈呈弯曲状,干扰了脂分子间范德华力的相互作用,所以排列比较疏松,增加 了膜的流动性。因此,脂质双分子层中含不饱和脂肪酸越多,膜的相变温度越低,其流动性也越大。 (2)脂肪酸链的长短:脂肪酸链的长短与膜的流动性有关。脂肪酸链短的相变温度低,膜流动性 增加:脂肪酸链长的相变温度高,膜流动性降低。这是因为脂肪酸链越短则尾端越不易发生相互作用, 在相变温度以下,不易发生凝集而增加了流动性:长链尾端之间不仅可以在同一分子层内相互作用,而 且可以与另一分子层中的长链尾端作用,使膜的流动性降低 (3)胆固醇的双重调节作用:动物细胞膜含较多的胆固醇,对膜的流动性具有双重调节作用,不同 的调节作用与胆固醇在不同温度时细胞膜中的定位相关。当温度在相变温度以上时,由于胆固醇的 醇环与磷脂分子靠近极性头部的烃链部分相结合,可以稳定质膜,减少流动性。而当温度在相变温度以 下时,由于胆固醇位于磷脂分子之间隔开了磷脂分子,可以有效防止脂肪酸链相互凝集,干扰晶态形成。 增加膜的流动性,因此,动物细胞膜中所含胆固醇的量可以有效地防止低温时膜流动性的突然降低 (4)卵磷脂与鞘磷脂的比值:哺乳动物细胞中,卵磷脂和鞘磷脂的含量约占膜脂的50%,其中卵磷 脂的脂肪酸链不饱和程度高,相变温度较低:鞘磷脂则相反,其脂肪酸链饱和程度高,具有较长的饱和脂 肪酸链,由于鞘磷脂分子间的作用力较强,相变温度也高,并且鞘磷脂的粘度比卵磷脂高6倍。因此,卵 358

第十五章细胞膜与细胞表面 磷脂与鞘磷脂的比值高则膜流动性增加:比值低则膜流动性减少。在细胞衰老过程中,细胞膜中卵磷脂 与鞘磷脂的比值逐渐下降,其流动性也随之降低。 (5)膜蛋白的影响:细胞膜上的膜脂结合蛋白质后对膜的流动性有直接影响。对于膜内在蛋白而 言,一旦膜蛋白嵌入膜脂疏水区后,使周围的脂类分子不能单独活动而形成界面脂(嵌入蛋白与周围脂 类分子结合而形成),嵌人的蛋白越多,界面脂就越多,膜的流动性越小。另一方面,在含有较多跨膜 白的细胞膜中.存在由骑草蛋白分割句围的宫脂风(iDid-rich region).磷胎分子只能在一个富脂区内目 由扩散,而不能扩散到邻近的富脂区。除上述原因外,膜脂的极性集团、环境温度、pH、离子强度等因素 也可以对膜脂的流动性产生一定的影响。如环境温度越高,膜脂流动性越大;在相变温度内,每下降 10℃,膜的黏性增加3倍,因而膜的流动性降低 3.膜蛋白的流动性膜蛋白的运动方式一般分为两类:侧向扩散和旋转运动。这两种分子运动方 式与膜脂分子相似,但移动速度较慢。 (1)侧向扩散:许多实验证明,膜蛋白在膜脂中可 莫蛋 以自由漂浮和在膜表面扩散。1970年,霍普金斯大学的 La四Fye和Michael Edidin用细胞融合结合免疫荧光 技术证明了膜蛋白的侧向扩矿散。他们首先将人和鼠的细 胞草用红鱼和绿鱼两种荧光抗体讲行标记然后将带有红 色和绿色两种不同荧光的细胞进行融合,起初融合的杂种 细胞一侧呈绿色荧光,一侧呈红色荧光。37℃继续培养 段时间后,研究中发现红色和绿色两种颜色的荧光在整个 杂交细胞膜上均匀分布,说明与荧光抗体结合的膜抗原蛋 白在膜平面内经侧向扩散运动而重新分布(图15-12)。 (2)旋转运动(rotation):或称旋转扩散(rotationa diffusion),膜蛋白能围绕与膜平面相垂直的轴进行旋转 蛋白抗你 运动。研究方向膜蛋白旋转扩散的速度比侧向扩散更 为缓慢。不同膜蛋白旋转速率也有很大差异,这与其分 子结物及所外不同的微环墙有关 7℃解育40分结 研究发现,细胞膜上并不是所有的膜蛋白都能自由 运动,有些细胞只有部分膜蛋白处于流动状态。膜蛋白 在脂质双层中的运动还受到许多其他因素的影响:整合 膜蛋白扩散 蛋白与周边蛋白的相互作用:膜蛋白与细胞骨架成分的 连接以及与膜脂的相互作用:若膜蛋白聚集形成复合 物,使其运动减慢;这些都会限制膜蛋白的运动性。如 果用细胞松驰素B处理细胞,阻断微丝的形成,可使膜 图15-12小鼠-人细胞融合过程中膜蛋白的 侧向扩散示意图 蛋白流动性增强 质膜的流动性是保证其正常功能的以要条件,且有十分重要的生理意义,例如跨膜物质运输、细胞 信息传递、细胞识别、细胞免疫、细胞分化以及激素的作用等都与膜的流动性密切相关。生物膜各种功 能的完成是在膜的流动状态下进行的,若膜的流动性降低,低于一定的阈值时,细胞膜固化、黏度增大到 一定程度时,许多酶的活动和跨膜运输将停止,膜内酶活性丧失,代谢终止:反之如果膜流动性过高,又 会造成膜的溶解。 三、细胞膜的分子结构模型 前面已经介绍了细胞膜的化学组成,但是细胞膜中膜脂和膜蛋白是如何排列并组建起来的?它们 之间的相互作用又是怎样的?这些对阐明细胞膜的功能活动是十分重要的。 350