
第十五章细胞膜与细胞表面 学习目标 通过本章学习,你应该能够 掌握细胞膜结构组成、物质运输功能与细胞连接、细胞黏附类型、结构和组成。 熟悉细胞膜的各种结构模型、细胞连接、细胞黏附结构和功能。 了解细胞膜与细胞表面的关系。 细胞膜(cell membrane),又称质膜(plasma membrane),是包围在细胞表面的一层薄膜,它将细胞中 的生命物质与外界环境分隔开,维持细胞特有的内环境。除质膜外,真核细胞内还有丰富的膜性结构, 它们组成了具有各种特定功能的膜性细胞器,例如内质网、高尔基复合体,溶酶体、线粒体等,组成这些 细胞器的膜统称内膜(endomembrane),内膜与细胞膜统称为生物膜(biomembrane),这些膜在化学组成、 分子结构和基本功能方面具有很多共性。 细胞膜不仅为细胞的生命活动提供了稳定的内环境,还行使着物质运输、信号传递、细胞识别等多 种复杂功能,并且与生命科学中的许多基本问题,如细胞增殖、分化、细胞识别黏附、代谢、能量转换等密 切相关,是细胞之间、细胞与细胞外环境之间相互交流的重要通道。 第一节细胞膜的结构与组成 细胞膜主要由脂类、蛋白质和糖类三种成分组成。一般而言,脂类重量约占50%,蛋白质约占 40%,糖类占1%~10%,几种成分所占比重会随着膜种类的不同而发生变化。细胞膜中脂类排列成双 分子层,组成膜的基本结构,形成了对水溶性分子相对不通透的屏障;蛋白质以不同方式与脂类结合,构 成膜的功能主体;糖类多分布于膜外表面,通过共价键与膜的某些脂类或蛋白质结合形成糖脂或糖蛋 白。此外,细胞膜中还含有少量水分、无机盐与金属离子等。 一、细胞膜的化学组成 (一)膜脂 细胞膜上的脂类称为膜脂(membrane lipid),约占膜成分的50%,一个动物细胞的质膜中大约含有 10个脂膜分子,它们主要有三种类型:磷脂、胆固醇和糖脂,其中以磷脂含量为最多(图151)。 1.磷脂大多数膜脂分子中都含有磷酸基团,被称为磷脂(phospholipid),占膜脂的50%以上,是 组成生物膜的主要成分。磷脂包括甘油磷脂(lycerophosphatide)和鞘磷脂(sphingomyelin,SM)两大类 甘油磷脂主要包括磷脂酰胆碱(卵磷脂)(phosphatidylcholine,PC)、磷脂酰乙醇胺(脑磷脂)(phosphati- dylethanolamine,PE)和磷脂酰丝氨酸(phosphatidylserine,PS)。此外,还有一种磷脂是磷脂酰肌醇(phos 350

第十五章细胞膜与细胞表面 蛋 镶蛋白 图151细胞膜的结构示意图 phatidylinosital,P),位于质膜的内层,在膜结构中含量很少,但在细胞信号转导中起重要作用。 甘油磷脂都以甘油为骨架,其磷酸基团分别与胆碱、乙醇胺、丝氨酸或者肌醇结合,即形成上述四种 类型磷脂分子。这些亲水的小基团在分子的末端与带负电的磷脂基团一起形成高度水溶性的结构域, 极性很强,被称为头部基团(head group)或亲水头部。磷脂中的脂肪酸链长短不一,且脂肪酸链是硫水 的,无极性,称为疏水尾部。因此,磷脂分子具有亲水头部和疏水尾部,被称为两亲性分子或兼性分子 (amphipathic molecule)。鞘磷脂是细胞膜上唯一不以甘油为骨架的磷脂,在膜中含量较少,但是在神经 元细胞膜中含量较多。它以鞘氨醇代替甘油,长链的不饱和脂肪酸结合在鞘氨醇的氨基上。目前研究 发现,鞘磷脂及其代谢产物神经酰胺、鞘氨酸及1-磷脂鞘氨酸参与各种细胞活动,如细胞增殖、分化和调 亡等(图15-2) 2.胆固醇胆固醇(cholesterol),是细胞膜中的另一类重要的脂类,分子较小,散布在磷脂分子之 间,也属于两亲性分子。其极性头部为羟基,靠近相邻磷脂分子的极性头部;中间为固醇环结构,连接 条短的疏水性烃链尾部,埋在脂双层的中央(图153)。 胆固醇分子对调节膜的流动性、加强膜的稳定性具有重要作用。膜上胆固醇分子的亲水性末端朝 向脂双层的外表面,而大部分结构嵌入磷脂的脂肪酸尾部中。与大多数膜脂不同,胆固醇在脂双层间倾 向于均匀分布(图15-4)。不同生物膜由各自特殊的脂类组成,如哺乳动物细胞膜上富含胆固醇和糖 脂而线粒体膜内富含心、赚指等 3.糖脂糖脂(lycolipid)由脂类和寡糖构成,其含量占膜脂总量的5%以下,在神经细胞膜上糖脂 含量较高,占5%~10%。糖脂也属于两亲性分子。动物细胞膜中的糖脂结构与鞘磷脂很相似,只是由 个或多个糖残基代替了磷脂酰胆碱而与鞘氨醇的羟基结合。如脑苷脂、神经节苷脂。糖脂普遍存在 于原核和真核细胞膜表面,位于膜的非胞质面单层,并将糖基暴露在细胞表面,其作用可能作为某些大 分子的受体,与细胞识别及信息传导有关。 目前已发现40余种糖脂,它们的主要区别在于其极性头部不同,由1个或几个糖残基构成。最简 单的糖脂是脑苷脂,其极性头部仅有一个半乳糖(Gl)或葡萄糖(G©)残基,它是髓鞘中的主要糖脂。 比较复杂的是神经节苷脂,其极性头部除含有半乳糖和葡萄糖外,还含有数目不等的唾液酸(也叫做N 乙酰神经氨酸,NANA)。神经节苷脂在神经元的质膜中最为丰富,占总脂类的5%~10%,但在其他类 型中的细胞中含量很少,迄今已鉴定了40多种神经节苷脂(图15-5)。所有细胞中,糖脂均位于质膜非 胞质表面,糖基暴露于细胞表面,其作用可能作为细胞表面受体,与细胞识别、黏附及信号传导有关。 膜脂都是双亲媒性分子,由于极性头部能与水分子形成氢键或静电作用溶于水,非极性尾部能与水 分子相互作用而疏水。所以当这些脂质分子被水环境包围时,它们就自发聚集起来,使疏水的尾部埋在 351
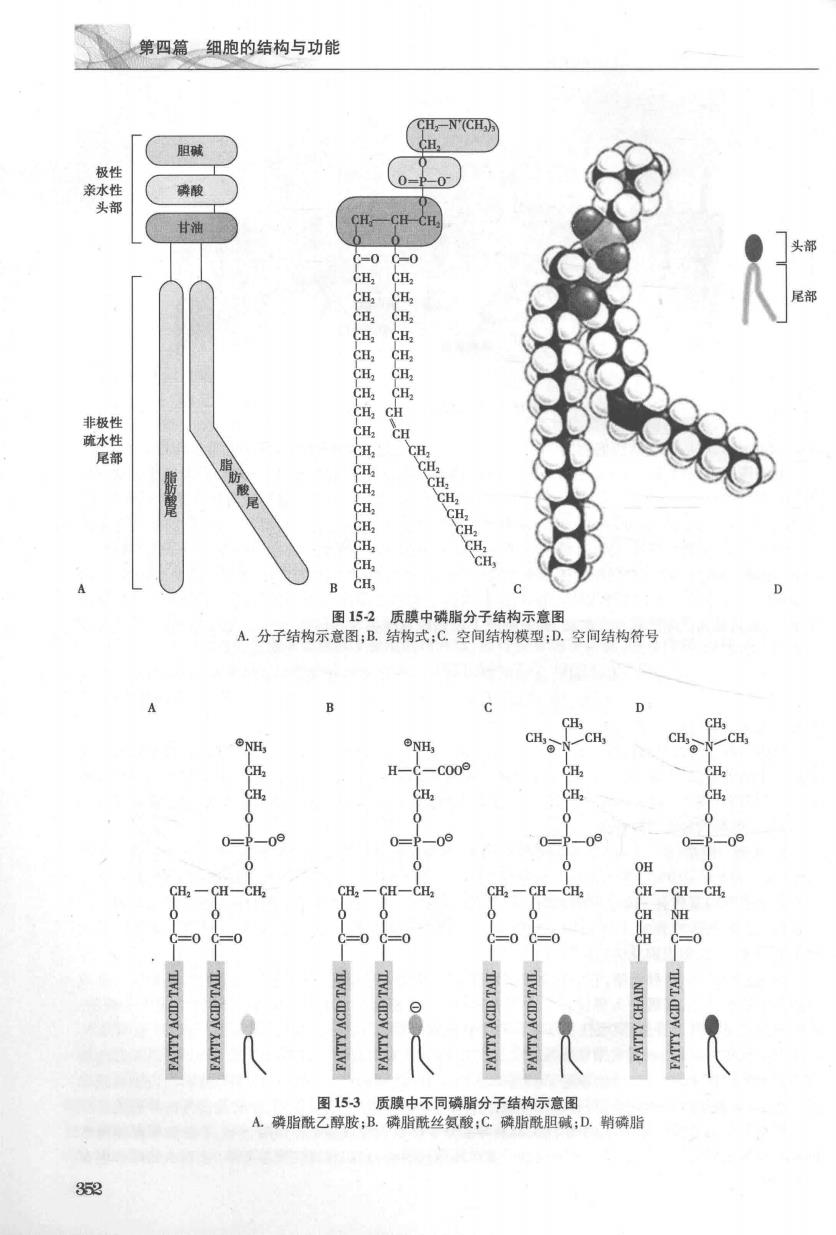
】第西草的结构与功能 CH一N(CH 胆碱 酸 甘油 CH =0 图152质膜中磷脂分子结构示意图 A.分子结构示意图:B.结构式:C空间结构模型:D.空间结构符号 H 图153质膜中不同脂分子结构示意图 A.南脂酰乙醇胺;B磷脂酰丝氨酸:C磷脂酰胆碱;D.鞘磷脂
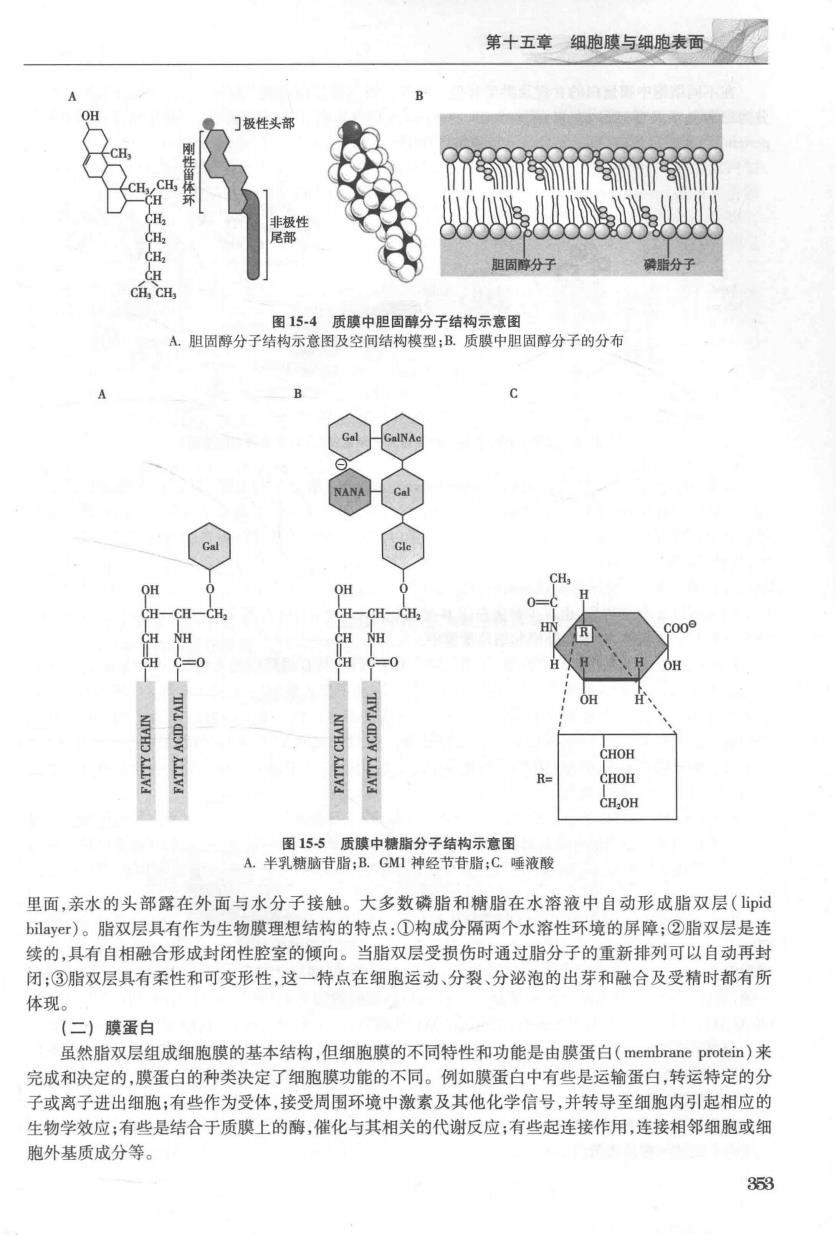
第十五章细胞膜与细胞表面 朝固障分子 分子 图154 A胆固醇分子 结构模型:B.质膜中胆固醇分子的分布 CH, CH CH 1=0 =0 CHOH CHOH CH.OH 图155质障中续脂分子结物示意图 A半乳糖脑苷脂:B,CM1神经节苷脂:C唾液酸 里面,亲水的头部露在外面与水分子接触。大多数磷脂和糖脂在水溶液中自动形成脂双层(pd bilay©r)。脂双层具有作为生物膜理想结构的特点:①构成分隔两个水溶性环境的屏障:②脂双层是连 续的,具有自相融合形成封闭性腔室的倾向。当脂双层受损伤时通过脂分子的重新排列可以自动再封 闭:③脂双层具有柔性和可变形性,这一特点在细胞运动、分裂、分泌泡的出芽和融合及受精时都有所 体现 (二)膜蛋白 虽然脂双层组成细胞膜的基本结构,但细胞膜的不同特性和功能是由膜蛋白(membrane protein)来 完成和决定的,膜蛋白的种类决定了细胞膜功能的不同。例如膜蛋白中有些是运输蛋白,转运特定的分 子或离子进出细胞;有些作为受体,接受周围环境中激素及其他化学信号,并转导至细胞内引起相应的 生物学效应:有些是结合于质膜上的酶,催化与其相关的代谢反应:有些起连接作用,连接相邻细胞或细 胞外基质成分等。 358

第四篇细胞的结构与功能 在不同细胞中膜蛋白的含量及类型有很大差异。根据膜蛋白与脂双层结合的方式不同,膜蛋白可 分为三种基本类型:膜内在蛋白(intrinsic protein)或整合蛋白(integral protein)、膜外在蛋白(extrinsic protein)和脂锚定蛋白(lipid anchored protein)(图15-6)。 H CYTOSO © E 28 图15-6膜蛋白在细胞膜中的结合方式示意图(示几种膜蛋白的类型) L.膜内在蛋白又称跨膜蛋白(transmembrane protein)整合蛋白或镶嵌蛋白,占膜蛋白总量的 70%一80%,也是两亲性分子,疏水部分位于脂双层内部,亲水部分位于脂双层外部。膜内在蛋白分为 单次跨膜、多次跨膜(图15-6A,B)和多亚基穿膜蛋白三种类型。单次穿膜蛋白的肽链只穿过膜双层 次,穿膜区一般含有20~30个疏水性氨基酸残基,以单条α螺旋构象穿越脂双层的疏水区。多次穿膜 蛋白含有多个由疏水性氨基酸残基组成的穿膜序列,通过数条α螺旋构象穿过脂双层。大多数穿膜蛋 白穿膜域都是a螺旋构象,也有少数蛋白以B-片层(B-pleated sheet)构象穿膜(图15-6C),这类穿膜蛋 白主要存在于线粒体、叶绿体外模和细菌质膜中。 膜内在蛋白由于存在疏水结构域,与膜的结合非常紧密,只有用强烈的去垢剂(detergent)才能从膜 上洗涤下来,如离子型去垢剂十二烷基磺酸钠(SDS),非离子型去垢剂Ttom-XIO0等。SDS为常用的离 子型去垢剂,它是两亲性的脂质样分子,含有一个带电的亲水区和疏水区(烃链)。由于SDS的极性端 带电荷(离子型)更易溶于水,所以形成了去垢剂-蛋白质复合物进入溶液中而将膜蛋白分离出来。SDS 对蛋白质的作用较强烈,能使蛋白质解析折叠引起变性,不利于对其进行功能研究。为获得有功能的膜 蛋白,可采用非离子型去垢剂。 Triton X-100是非离子型去垢剂,它的极性端不带电荷,可使细胞膜崩解,但对蛋白质的作用比较温 收音5华生发网 出去。 2.膜外在蛋白又称为周边蛋白(peripheral protein),占膜蛋白总量的20%-30%。是一类与细胞 膜结合比较松散的不插入脂双层的蛋白质,分布在质膜的胞质侧或胞外侧。一些周边蛋白通过非共价 键附着在脂类分子头部极性区域或穿膜蛋白亲水区的一侧,间接与膜结合;一些周边蛋白位于膜的胞质 一侧,通过暴露于蛋白质表面的α螺旋的疏水面与脂双层的胞质面单层相互作用面与膜结合(图15 6D,G,H)。周边蛋白为水溶性蛋白,它与膜的结合比较弱,一般使用一些温和的方法,就可以将它们从 膜上分离下来,而不需要破坏膜的基本结构,常用的方法有改变溶液的离子强度或浓度、甚至提高温度 就可以从膜上分离下来外周蛋白。有时很难区分整合蛋白和外周蛋白,主要是因为一个蛋白质可以由 多个亚基构成,有的亚基为跨膜蛋白,有的则结合在膜的外部。 周边蛋白与细胞膜之间通常有一种动态关系,根据功能的需要募集到膜上或者从膜上释放出去。 质膜内表面的一些周边蛋白有的作为酶或者传递穿膜信号的因子发挥作用,与质膜外表面相连的一些 354