
第十二章RNA的生物合成 学习目标 通过本章的学习,你应该能够 掌握转录的概念及其反应体系,包括模板、原料、酶及其化学反应。启动子的 概念与作用。掌握真核生物mRNA的加工修饰方式包括前体mRNA的5 端加帽3'端加尾、剪接、mRNA的选择性剪接和RNA编辑等。 熟悉复制与转录的区别。不对称转录、模板链、编码链等基本概念。σ亚基的 作用。原核基因启动子的结构。hRNA、外显子、内含子、断裂基因等 概念。 了解RNA生物合成的主要方式。大肠杆菌RNA聚合酶的组成。真核生物 RNA聚合酶的分类和作用。真核基因启动子的特征。原核生物转录与真 核生物转录的基本过程,以及两者的异同。原核生物mRNA的加工。 RNA和RNA的加工修饰方式。RNA的自我剪接和催化功能。RNA 复制。 在生物界,RNA的合成有两种方式,包括转录(transcription)和RNA复制(RNA replication)。转录 是DNA指导的RNA合成过程,即以DNA为模板合成RNA的过程,也就是把DNA的核苷酸序列(原 件)“抄录”为RNA的核苷酸序列(副本或抄本),这是生物体内NA合成的主要方式。本章主要介绍 转录。RNA复制是RNA指导的RNA合成过程,即以RNA为模板合成RNA的过程,常见于以RNA为 遗传物质的RNA病毒。转录是基因表达为蛋白质产物的首要步骤,转录的产物—RNA包括mRNA RNA和RNA等,其中mRNA把遗传信息从染色体内产存的状态“妙录"出来,作为蛋白质合成的直接 模板,而RNA和RNA不用作蛋白质合成的模板,但通过转运氨基酸和组成核糖体等机制参与蛋白质 的生物合成(见第十三章)。 在遗传信息传递过程中,转录和复制是两种不同的生物学过程,两者既有不同,也有诸多相似之处 (表12-1)。 表121复制和转录的比较 复制 转录 定义 以DNA为模板合成DNA的过程 以DNA为模板合成RNA的过程 相同点 ①都是酶促的核苷酸聚合过程: ⑤核苷酸之问都以磷酸二酯键相连; ②都是以DNA为模板; ⑥都服从碱基配对规则: ③都是以核苷酸为原料: ⑦产物都是很长的多核苷酸链; ④合成方向都是5'→3': 278

第三篇遗传信息传递及其调控 续表 复制 转录 不同点 ①模板:DNA两股单链都可作为模板 仅一股DNA单链即模板链被转录 ②原料:四种dNTP(dATP,dGTP,dCTP,dTP) 四种NTP(ATP、GTP、CTP、UTP) 3记对:G=C、A=1 G=C,A=T、A=0 ④磷:DNA聚合酶、拓扑异构酶,DNA连接酶等 RNA聚合酵 ⑤产物:子代双链DNA mRNA、tRNA,rRNA等 ⑥引物:需要短RNA片段作为引物 不需要引物 ⑦方式:半保留复制、双向复制、半不连续复制 不对称转录 第一节转录的概念及其反应体系 转录是以DNA双链中的一股单链作为转录模板,以四种NTP为原料,按照碱基配对原则,由RNA 聚合酶(RNA polymerase,RNA pol)催化合成RNA的过程。转录属于酶促反应,转录的反应体系包括 DNA模板4种NTP原料、RNA聚合酶和一些蛋白质因子等。 一、转录的模板 复制和转录的模板都是D八A,但因为两者的目的截然不同,故在模板的利用方式上也截然不同 复制是为了保留物种的全部遗传信息,所以,基因组DNA全长均需复制,并且在每次细胞分裂时进行且 只进行一次,以确保将一份完整的遗传信息传递给子代。然而,转录是为了将基因组DNA中的功能性 遗传信息抄录出来,表达为有特定功能的RNA或蛋白质分子,所以转录是有选择性地抄录基因组DNA 的特定部分,是不连续的。在不同类型的细胞中、不同的生理或病理条件下,被转录的基因以及转录的 水平或效率高低明显不同,可以说是“按需选择性转录”。另一方面,在庞大的细胞基因组DNA链上 也并非任何区段都可以被转录,尤其对于真核生物基因组D八A来讲,有不少区域是不被转录的。而对 于被转录的DNA分子双链来讲,也只有一股链作为模板指导转录,另一股链则不被转录,而且不同基因 转录时使用的模板链并非总是在同一单链上。转录对DNA双链的这种选择性有时被特称为不对称转 录(asymmetric transcription)(图12-1)。 DNA双链中,转录时直接作为模板按照碱基配对规律指导RNA合成的一股单链,称为模板链或 Watson链(templatestrand)。相对的另 一股单链虽不作为转录的直接模板,但因其序列与模板链序列互 补,故其碱基序列与新合成的RNA链一致(只是T被U取代),也就是说新合成的RNA链实际上抄录 了这条链的序列。若转录产物是RNA,则可用作蛋白质翻译的模板,按照遗传密码规则讲一步决定所 合成蛋白质的氨基酸序列,所以与模板链相对应这股单链就被称为编码链或Cnck链(coding strand),也 称正义链或有意义链(sense strand),(图l2-2)。相应地,模板链则又可称为负义链或反义链(antisense sand)。转录总是从5→3'方向进行RNA合成,所以转录总是沿着模板链3'一→5'方向进行。文献刊出 的DNA序列,为避免繁琐和便于查对遗传密码,一般只写出编码链。 5,GCATTGAATCTGGTC.3编 基因A 3 CGTAACTTAGACCAG.S模板链 基因B 基因C 基因D DNA 5 5.GCAUUGAAUCUGGUC.3'mRNA 潮译 一模板链→转录方向 N.Ala Leu Aan Leu Val.C.多肽迹 图121一段染色体DNA上不同基因的转录方向 图12-2DNA双链的模板链与编码链 2
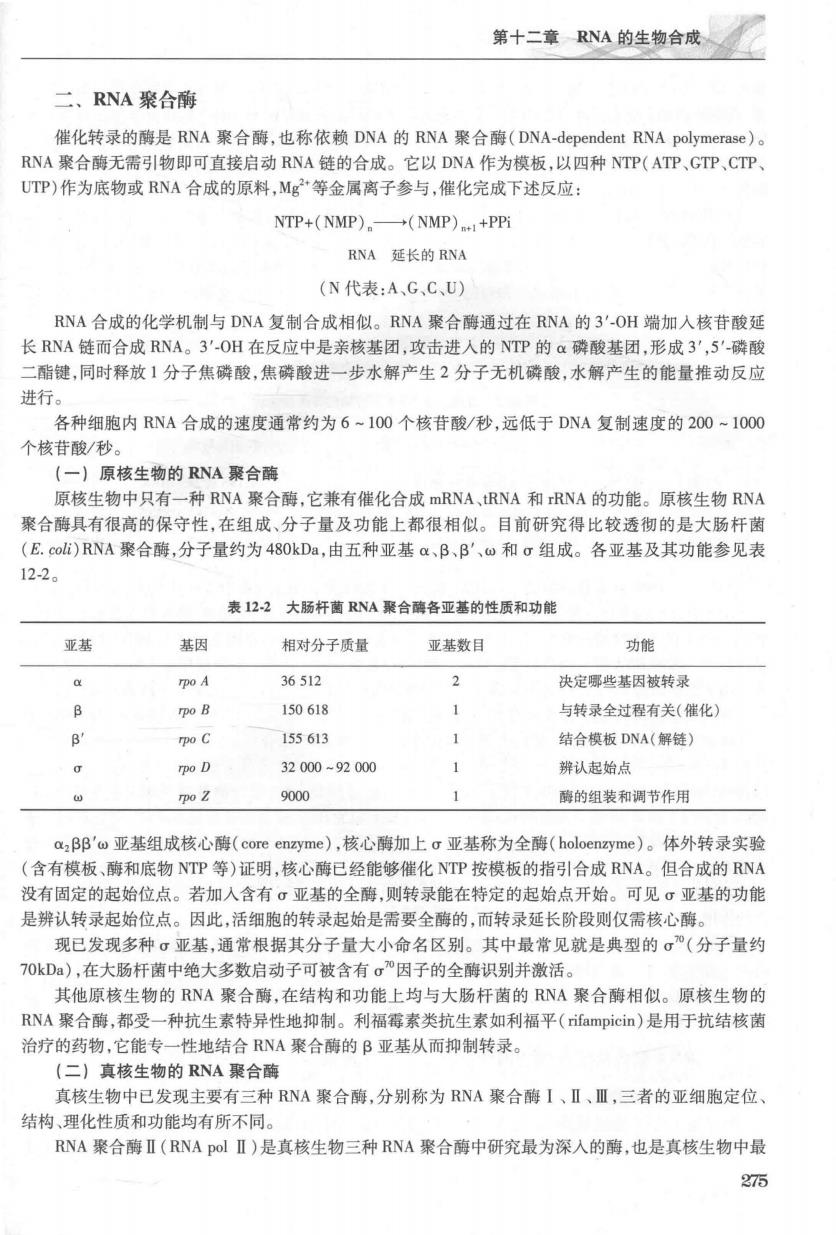
第十二章RNA的生物合成看 二、RNA聚合酶 催化转录的酶是RNA聚合酶,也称依赖DNA的RNA聚合酶(DNA-dependent RNA polymerase) RNA聚合酶无需引物即可直接启动RNA链的合成。它以DNA作为模板,以四种NTP(ATP、GTP,CTP UTP)作为底物或RNA合成的原料,Mg2·等金属离子参与,催化完成下述反应: NTP+(NMP).一→(NMP)1+PP RNA延长的RNA (N代表:A,GC、U) RNA合成的化学机制与DNA复制合成相似。RNA聚合德通过在RNA的3'-OH端加人核苷酸延 长RNA链而合成RNA。3'OH在反应中是亲核基团,攻击进人的NTP的Q磷酸基团,形成3',5'磷酸 二酯键,同时释放1分子焦磷酸,焦磷酸进一步水解产生2分子无机磷酸,水解产生的能量推动反应 进行。 各种细胞内RNA合成的速度通常约为6~1O0个核苷酸/秒,远低于DNA复制速度的200-1000 个核苷酸/秒。 【一】原核生物的RNA襄合随 原核生物中只有一种RNA聚合酶,它兼有催化合成mRNA,tRNA和rRNA的功能。原核生物RNA 聚合酶具有很高的保守性,在组成、分子量及功能上都很相似。目前研究得比较透彻的是大肠杆菌 (E.coli)RNA聚合酶,分子量约为48OkDa,由五种亚基a,B、B'、和组成。各亚基及其功能参见表 12-2。 表122大肠杆药RNA聚合砖各亚基的性质和功能 亚基 基因 相对分子质量 亚基数目 功能 36512 2 决定哪些基因被转录 B rpo B 150618 与转录全过程有关(催化】 B' o C 155613 1 结合模板DNA(解链) g po D 32000-92000 1 辨认起始点 rpo Z 9000 1 酶的组装和调节作用 a,BB'w亚基组成核心酶(core enzyme),核心酶加上g亚基称为全酶(holoenzyme)。体外转录实验 (含有模板、酶和底物NTP等)证明,核心酶已经能够催化NTP按模板的指引合成RNA。但合成的RNA 没有固定的起始位点。若加入含有σ亚基的全酶,则转录能在特定的起始点开始。可见亚基的功能 是辨认转录起始位点。因此,活细胞的转录起始是需要全酶的,而转录延长阶段则仅需核心酶。 现已发现多种σ亚基,通常根据其分子量大小命名区别。其中最常见就是典型的σ”(分子量约 70kDa),在大肠杆菌中绝大多数启动子可被含有。”因子的全酶识别并激活 其他原核生物的RNA聚合酶,在结构和功能上均与大肠杆菌的RNA聚合酶相似。原核生物的 RNA聚合酶,都受一种抗生素特异性地抑制。利福霉素类抗生素如利福平(rifampicin)是用于抗结核菌 治疗的药物,它能专一性地结合RNA聚合酶的B亚基从而抑制转录。 (二】直核生物的RNA聚合燕 真核生物中已发现主要有三种RNA聚合酶,分别称为RNA聚合酶I、Ⅱ、Ⅲ,三者的亚细胞定位、 结构、理化性质和功能均有所不同。 RNA聚合酶Ⅱ(RNA polⅡ)是真核生物三种RNA聚合酶中研究最为深入的酶,也是真核生物中最 275
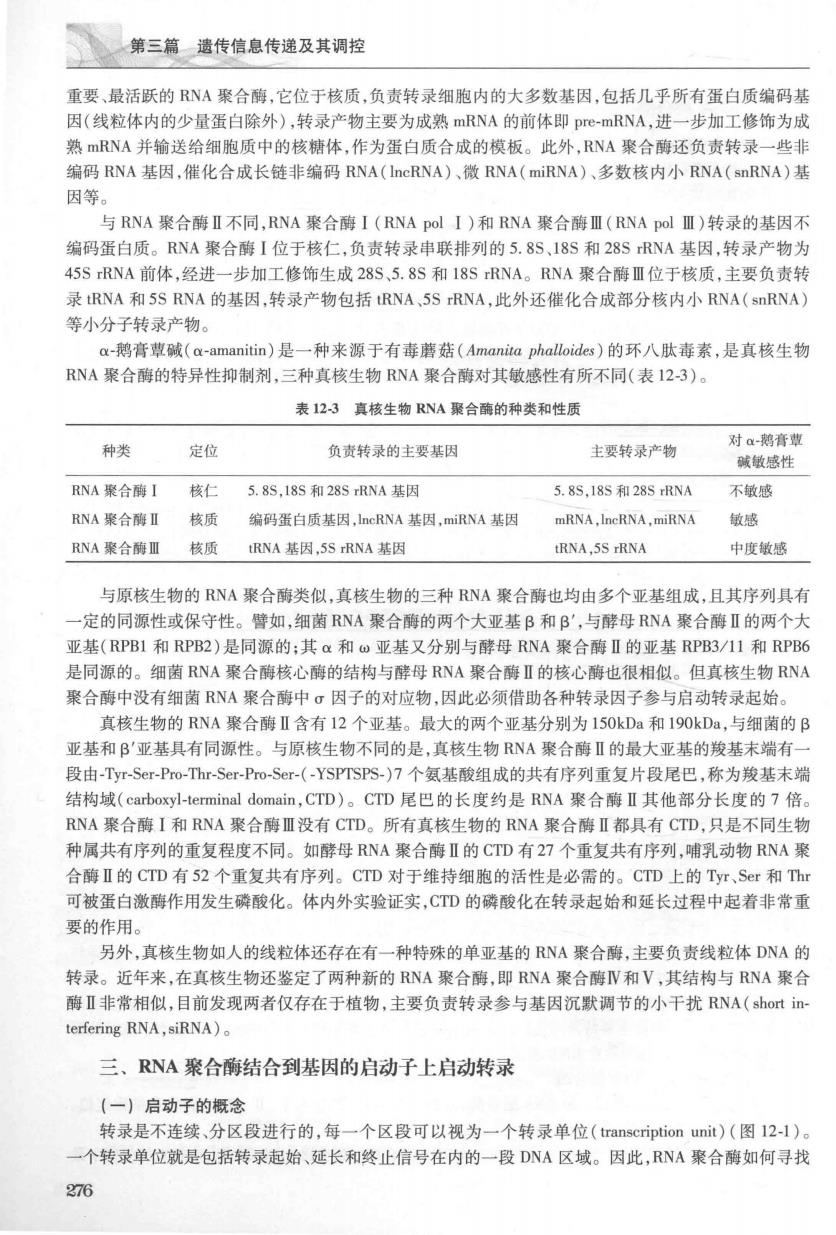
第三篇遗传信息传递及其调控 重要、最活跃的RNA聚合酶,它位于核质,负责转录细胞内的大多数基因,包括几乎所有蛋白质编码基 因(线粒体内的少量蛋白除外),转录产物主要为成熟mRNA的前体即pre-mRNA,进一步加工修饰为成 熟mRNA并输送给细胞质中的核糖体,作为蛋白质合成的模板。此外,RNA聚合酶还负责转录一些非 编码RNA基因,催化合成长特非绾码RNA(DcrNA),微RNA(mirNA),多数核内小RNA(BORNA)其 因等 与RNA聚合曹Ⅱ不同,RNA聚合酶I(RNA pol I)和RNA聚合酶Ⅲ(RNA polⅢ)转录的基因不 编码蛋白质。RNA聚合酶I位于核仁,负责转录串联排列的5.8S,18S和28SRNA基因,转录产物为 45 S IRNA前体.经讲一步加工修饰生成28S,5.8S和18SRNA。RNA聚合德Ⅲ位干核质.主要负青转 录RNA和5S RNA的基因,转录产物包括RNA,5SRNA,此外还催化合成部分核内小RNA(snRNA 等小分子转录产物。 ax-鹅膏碱(a-amanitin)是一种来源于有毒蘑菇(Amanita phalloides)的环八肽毒素,是真核生物 RNA聚合酶的特异性抑制剂,三种真核生物RNA聚合酵对其敏感性有所不同(表12-3)。 表123真核生物RNA聚合酶的种类和性质 种类 对a-鹅膏型 定 负责转录的主要基因 主要转录产物 碱敏感性 RNA聚合酶I 核 5.8S,18S和28SRNA基因 5.8S,18S和28SRNA 不敏感 RNA聚合酶I 核质 编码蛋白质基因,neRNA基因,miRNA基因 mRNA,IneRNA,miRNA 敏感 RNA聚合酶Ⅲ 核质 tRNA基因,5 S IRNA基因 tRNA.5S rRNA 中度敏感 与原核生物的RNA聚合酶类似,真核生物的三种RNA聚合酶也均由多个亚基组成,且其序列具有 -定的同原性或保守性。昏如,细菌RNA聚合酶的两个大亚基B和B',与酵母RNA聚合特Ⅱ的两个大 亚基(RPB1和RPB2)是同源的:其a和亚基又分别与酵母RNA聚合酶Ⅱ的亚基RPB3/I1和RPB6 是同原的。细菌RNA聚合酶核心酶的结构与酵母RNA聚合酶Ⅱ的核心酶也很相似。但直核生物RNA 聚合酶中没有细菌RNA聚合酶中σ因子的对应物,因此必须借助各种转录因子参与启动转录起始 真核生物的RNA聚合聘Ⅱ含有12个亚基。最大的两个亚基分别为150kDa和190kDa,与细菌的B 亚基和B'亚基具有同源性。与原核生物不同的是,真核生物RNA聚合酶Ⅱ的最大亚基的羧基末端有一 段由-TyT-Ser-Pro-Thr-Ser-Po-Ser-(~YSPTSPS-)7个氨基酸组成的共有序列重复片段尾巴,称为羧基末端 结构域(carboxyl-terminal domain,CTD)。CTD尾巴的长度约是RNA聚合酶Ⅱ其他部分长度的7倍。 RNA聚合酶I和RNA聚合酶Ⅲ没有CTD。所有真核生物的RNA聚合酶Ⅱ都具有CTD,只是不同生物 种属共有序列的重复程度不同。如酵母RNA聚合酶Ⅱ的CTD有27个重复共有序列,哺乳动物RNA聚 合酶Ⅱ的CTD有52个重复共有序列。CTD对于维持细胞的活性是必需的。CTD上的Tyr、Ser和Th 可被蛋白激酶作用发生磷酸化。体内外实验证实,CTD的磷酸化在转录起始和延长过程中起着非常重 要的作用 另外,真核生物如人的线粒体还存在有一种特殊的单亚基的RNA聚合酶,主要负责线粒体DNA的 转录。近年来,在真核生物还鉴定了两种新的RNA聚合酶,即RNA聚合酶VN和V,其结构与RNA聚合 酶I非常相似,目前发现两者仅存在于植物,主要负责转录参与基因沉默调节的小干扰RNA(short in erfering RNA.siRNA) 三、RNA聚合酶结合到基因的启动子上启动转录 【一】启动子的据念 转录是不连续、分区段进行的,每一个区段可以视为一个转录单位(transcription unit)(图12-1) 一个转录单位就是包括转录起始、延长和终止信号在内的一段DNA区域。因此,RNA聚合酶如何寻找 376

第十二章RNA的生物合成 到每个待转录单位的起始位点是启动转录的关键。 DNA模板上转录开始的位点称为转录起始位点(transeription start site,TSS),方便起见,通常将转 起始位点的核苷酸或碱基位置设定为+1,上游和下游的核苷酸或碱基序数则相应地分别用负数和正数 标示。启动子(promoter)则是指通常位于基因转录起始位点上游、能够与RNA聚合酶和其他转录因子 结合并进而调节其下游目的基因转录起始和转录效率的一段DNA序列。因此,启动子是转录调控的关 键部位。 对启动子的研究,常采用一种巧妙的方法即RNA聚合酶保护法:先把一段基因分离出来,然后和损 纯的RNA聚合酶混合,再加进核酸外切酶作用一定时间后,DNA链受核酸外切商水解,生成游离核苷 酸。但是有一段4O~6Obp的DNA片段是完整的。这表明这段DNA因与RNA聚合酶结合而受到保 护。受保护的DNA位于结构基因的上游。所以这一被保护的DNA片段就是被RNA聚合酶辨认和识 别的区域,并在这里准备开始转录,该区域就是核心启动子区域。 因为原核生物和真核生物的NA聚合酶结构不同,所以两者的基因启动子的特征也有所不同,这 也决定了两者的转录过程和转录调控机制有所差异。 (二)原核生物基因启动子的特征 原核生物的绝大多数基因按照功能相关性成簇地串联排列于染色体上,共同组成一个转录单位,即 操纵子(operon)(见第14章)。也就是说,一个操纵子通常包括多个编码蛋白的结构基因及其上游的启 动子等调控序列(对于原核生物基因来讲,因启动子隶属于操纵子的一部分,故多又被称为启动序列) 对大量的原核生物基因操子中的启动序列讲行序列分析发现其几有明显的特征,含有高度保守 的共有序列(consensus sequence)。如图12-3所示,其典型的共有序列主要是位于转录起始位点(即+1) 上游-35bp处的TTGACA和-10bp处的TATAAT。-10区的共有序列是1975年由David Pribnow首次 发现,故又称为Pribnow盒(Pribnow box)。转录起始位点(+l)即第一个核苷酸通常为腺嘌呤核苷酸。 转录起始位点 35区 -108 操纵子 TTGACA TATAAT- m 图12-3典型的原核生物启动子DNA序列 一般来讲,启动子的相应序列与一致性序列战接近,其启动子活性械高,就能够在一定的时间内把 始合成更多的RNA,基因的转录水平或效率就越高。在一些转录水平非常高的RNA基因的启动子中 往往在其更上游的-40~-6Obp还存在有一种额外的能够与RNA聚合酶α亚基的羧基末端结构域结合 的UP(upstream promoter)元件,这就能进一步增强RNA聚合酶与启动子的结合而确保高水平的转录。 RNA聚合酶全酶中的σ因子分别通过其分子结构中的一段“螺旋转角螺旋”模体和另一段α蝴 旋区与待转录基因启动子区的-35区和-10区结合。σ因子与-35区的结合主要是简单地使RNA聚合 酶能相对固定地结合在启动子区,而。因子与-10区的结合则在启动转录方面起着尤为重要的作用, 如导致-10区附近的DNA双螺旋解开并使解开的单链固定进而启动转录(图124)。 277