
第十章物质代谢的联系与调节 学习目标 通过本章的学习,你应该能够: 掌握关键酶的概念和特点以及关键酶活性的调节方式(快速调节和迟缓调 节),快速调节和迟缓调节 熟悉能量代谢的相互联系与制约,三大营养物质(糖、脂、蛋白质)代谢的相互 联系,体内代谢调节的基本方式。 了解物质代谢的特点,重要组织器官的代谢特点及联系:两类激素的受体模 式:饱食、空腹、饥饿、应激状态下的整体物质代谢调节:代谢综合征的概 念,糖尿病的主要代谢改变:代谢组学的概念。 生物体的生存和健康有赖于其机体不断地与外界进行物质交换。食物中的糖、脂及蛋白质经消化 吸收进人体内,在细胞内进行分解代谢提供能量,以满足生命活动的需要:同时也可通过合成代谢转变 成机体自身的蛋白质、脂类、糖类,以参与机体的构成。机体这种和环境之间不断进行的物质交换,即物 质代谢。物质代谢是生命的本质特征,也是生命活动的物质基础。 第一节物质代谢的特点及相互联系 一、物质代谢的特点 (一)整体性和通用性 1.各物质代谢途径相互协调“汇聚”为整体体外摄人的营养物,经消化吸收进入机体后的代谢不 是彼此孤立,而是同时进行的,且彼此互相联系、转变以及依存,构成统一的整体。例如进食后,摄入体 内的葡萄糖增多。机体除加强糖原合成外,葡萄糖分解代谢也同时加强,以释放充足的能量保证糖原 脂肪、蛋白质等物质合成:同时糖原分解、脂肪动员及蛋白质分解过程受到抑制。 此外,内源性代射物(体内各组织细胞分解来的糖、脂、氨基酸)也通衬各物质的共同代射池参与机 体的整体性物质代谢。例如:无论是由食物消化吸收的氨基酸,还是由机体自身组织蛋白降解产生的氨 基酸,或者机体自身合成的营养非必需氨基酸,均混为一体构成氨基酸代谢池,参与各种组织的代谢。 因此,细胞内各种物质的共同代谢池是物质代谢形成整体性的基础 2.ATP是能量“流通”的共同形式ATP是能量的直接利用形式。糖、脂及蛋白质在体内氧化分解 释出的能量,大部分储存在ATP的高能磷酸键中。体内其他的高能化合物,如磷酸肌酸、其他形式的 NTP、琥珀酰辅酶A等均需要转化为ATP进行利用。生命活动所需的一切能量如物质合成,肌肉收缩, 235

第二篇物质代谢及其调节 神经冲动的传导等均直接由ATP提供。ATP犹如一种能量货币,是能量交换的媒介,整合了细胞中复 杂的能量循环。 3.NADPH是合成代谢所需的还原当量许多参与氧化分解代谢的脱氢酶常以NAD'为辅酶。而 在很多还原性生物合成途径中,产物比其前体更具有还原性,因此参与这类合成代谢的还原酶则多以 NADPH为轴酶,提供还原当量。例如,经磷酸戊糖途径生成的NADPH既可为乙酰辅酶A合成脂酸,又 可为合成阳固醇提供环原当量。坏原当最NADP日是联系氧化与还原反应,整合分解与合成代谢途径 的“桥柔”。 4.分解/合成代谢途径具有共同的中间产物各种生物分子合成所需的小分子物质大多来源于分 解代谢途径。很多分解代谢途径都具有共同的中间产物。例如,丙酮酸、草酰乙酸是联系糖代谢与氨基 酸代谢的枢纽分子。三羧酸循环既是糖、脂肪及氨基酸分解代谢的最终共同通路,也是协调、联系三大 物质公解及相互转化的椒纽机制。通常将既与合成又与分解有关的代射途径称为两用代谢涂径。中间 产物和两用代谢途径是整合各种代谢途径的必经之路。 5.线粒体是代谢途径和代谢调节信号的整合点线粒体不仅是氧化磷酸化生成ATP的场所,也是 两用代谢途径的“起源地”,是细胞各种区域分隔的中心,担负联系、分流、整合细胞各种代谢途径的功 能。此外,线粒体具有调节细胞内C浓度、各种活泼氧水平的功能,因此线粒体功能与细胞信号转导 代谢稳定的调节,以及细胞凋亡密切相关。线粒体是整合物质代谢的功能产所。 (二)代谢调节维持稳态 生物体的各种物质代谢错综复杂,以对抗外环境变化,维持内环境恒定,即稳态。从生物化学角度 认识稳态,就是机体通过精细的调节机制,不断调节各种物质代谢的强度、方向和速度,以补偿外环境变 化而维持的代谢动力学动态稳定状态 一代谢稳态(metabolic homeostasis)。而这种通过改变各种物质 代谢的强度、方向和速度,以对抗代谢产物浓度变化的机制即为代谢调节。代谢调节普遍存在于生物 界,是生物的重要特征,是生命在进化过程中逐步形成的一种适应能力。 (三)各组织、器官物质代谢各具特色 由于各组织、器官分化不同,所含酶系的种类和含量各有差异,因而形成各组织、器官各具特色的代 谢特点。例如肝是维持血糖水平恒定的重要器官,它不仅能进行糖异生,还能进行糖原的合成和分解 再如肝只能进行酮体的合成却不能利用酮体:肝糖原的分解可以作为血糖的补充来源,而肌糖原分解 却只能分解供能,而不能直接补充血糖。这种物质代谢的组织特异性,对理解有关疾病的生化机制十分 重要 (四)代谢途径具有多样性 体内的物质代谢通常是以一系列酶促反应组成。代谢途径有多种: L.直线途径一般指从起始物到终产物的整个反应过程中无代谢支路。如DNA的生物合成等 2.分支途径一般是指代谢物可通过某个共同中间产物进行代谢分途,产生两种或更多种产物 例如以6磷酸葡萄糖为分支点的糖代谢,6磷酸葡萄糖可以进人糖酵解途径,也可进入磷酸戊糖途径, 还可以进人糖原合成途径。再如糖酵解途径产生的丙酮酸,在相对缺氧时被还原为乳酸,在有氧条件下 则氧化脱羧生成乙酰辅酶A。 3.循环途径循环中的中间产物可以反复生成,反复利用,使生物体能经济高效地进行代谢变换 而且循环反应可以从任一中间产物起始或终止,可大大提高代谢变化的灵活性。例如三羧酸循环、鸟氨 酸循环 二、物质代谢的相互联系 (一)三大营养物在能量代谢上相互联系、相互制约 糖、脂及蛋白质/氨基酸均可在体内氧化分解释放能量。从能量供应的角度看,这三大营养物可以 互相代替,并互相制约。一般情况下,机体氧化分解供能以糖、脂为主,较少分解蛋白质/氨基酸供能。 236

第十章物质代谢的联系与调节 这是因为,人类普通膳食所含热量物质主要是糖类(占总热量的60%~70%)和脂肪(20%-25%):后 者又因其含水少便于储存,故成为机体储能的主要形式。与糖、脂不同,蛋白质虽然也能氧化分解提供 能量,但蛋白质是机体的“建筑材料”,其主要功能是参与细胞的构成,通常并无多余储存,故机体尽可 能节约蛋白质的消耗:再则,蛋白质/氨基酸氧化分解供能时产生氨(详见第7章),氨的转化可能会使 机体冒“损肝伤肾”风险而付出高昂代价。 虽然三大营养物在体内氧化分解的代谢途径各不相同,但它们的共同中间产物是乙酰CA,最终分 解机制是三羧酸循环,产生能量主要通过氧化磷酸化转化为ATP形式。所以任一供能物质分解代谢占 优势,常可抑制其他供能物质的分解。例如,脂肪分解增强、生成的ATP增多,ATP/ADP比值增高,可变 构抑制葡萄糖分解代谢中的限速酶一磷酸果糖激酶1活性,从而抑制葡萄糖分解代谢。相反,若供能物 质不足,体内ATP减少,ADP积存增多,则可变构激活磷酸果糖激酶-1,加速体内糖的分解代谢。 (二)物质代谢通过共同中间产物相互联系、相互转化 体内糖、脂、蛋白质/氨基酸和核酸等的代谢不是彼此独立,而是相互关联的。它们通过共同的中间 代谢物(即两条代谢途径汇合时的中间产物)和两用代谢途径(如三羧酸循环和生物氧化等)联成整体。 经此整合,三大营养物之间还可以互相转化:当一种物质代谢障碍时可引起其他物质代谢的紊乱,如 尿病时糖代谢的障碍,可引脂代谢、蛋白质代谢甚至水盐代谢的素乱。 1.糖代谢与脂代谢的相互联系 (1)糖可转变为脂肪:当从膳食中摄入的糖量超过体内能量消耗时,除合成少量糖原储存外,生成 的柠檬酸及ATP可变构激活乙酰辅酶A羧化酶,使由糖代谢而来的大量乙酰辅酶A得以羧化成丙二酰 铺酶A,进而合成脂酸及脂肪在脂肪组织中储存。这就是为什么摄取不含脂肪的高糖膳食可使人肥胖 及血甘油三酯升高的原因。 (2)脂肪中甘油部分可转变为糖:当大量脂酸分解时,在肝、肾、肠等组织中甘油激酶的作用下,将 甘油转变成3磷酸甘油,后者通过糖异生途径转变生成葡萄糖或糖原:但脂酸不能在体内转变为糖,因 为脂酸分解生成的乙酰辅酶A不能转变为丙酮酸。所以脂肪只有甘油部分可转变为糖。 (3)脂肪的分解代谢受糖代谢的影响:糖代谢的正常进行是脂肪分解代谢顺利进行的前提。因为 脂酸氧化的产物乙酰辅酶A必须与草酰乙酸缩合成柠檬酸后进入三羧酸循环,才能彻底氧化,而草酰 乙酸主要靠糖代谢产生的丙酮酸羧化生成。当饥饿、糖供给不足或糖代谢障碍时,脂肪作为主要的供能 物质大量动员,脂酸B氧化加强,由于缺乏糖代谢产生的草酰乙酸,乙酰辅酶A不能有效地进入三羧酸 循环而合成大量酮体,造成血酮体升高,产生高酮血症。 2.糖代谢与氨基酸代谢的相互联系 (1)绝大多数氨基酸可转变为葡萄糖:参与蛋白质构成的20种基本氨基酸,除生酮氨基酸(亮氨 酸、赖氨酸)外,都可通过脱氨作用生成相应的α酮酸。这些α-酮酸可转变成某些中间代谢物如丙酮 酸、草酰乙酸、α-酮戊二酸等,循糖异生途径转变为糖。如精氨酸、组氨酸及脯氨酸均可通过转变成谷氨 酸进一步脱氨生成α-酮戊二酸,经草酰乙酸转变成磷酸烯醇式丙酮酸,再循糖酵解逆行途径转变成糖。 (2)糖可以转变为非必需氨基酸:糖代谢的一些中间代谢物,如丙酮酸、α酮戊二酸、草酰乙酸等 也可氨基化生成某些非必需氨基酸。但苏、甲硫、赖、亮、异亮、缬、苯丙及色氨酸等8种必需氨基酸不能 由糖代谢中间物转变而来,必须由食物供给,所以这就是为什么食物中的糖不能替代蛋白质,而蛋白质 却能替代糖和脂肪供能的重要原因。 3.脂类代谢与氨基酸代谢的相互联系 (1)蛋白质可以转变为脂肪:氨基酸无论生糖、生酮(亮氨酸、赖氨酸)或生酮并生糖氨基酸(异亮、苯 丙、色、酪、苏氨酸)分解后均可生成乙酰辅酶A,后者可合成脂酸进而合成脂肪,即蛋白质可转变为脂肪。 (2)氨基酸可作为合成磷脂的原料:丝氨酸脱羧可变为胆胺,胆胺经甲基化可变为胆碱。丝氨酸 胆胺及胆碱分别是合成丝氨酸磷脂、脑磷脂及卵磷脂的原料。此外,乙酰辅酶A也可合成胆固醇以满 足机体的需要。 237
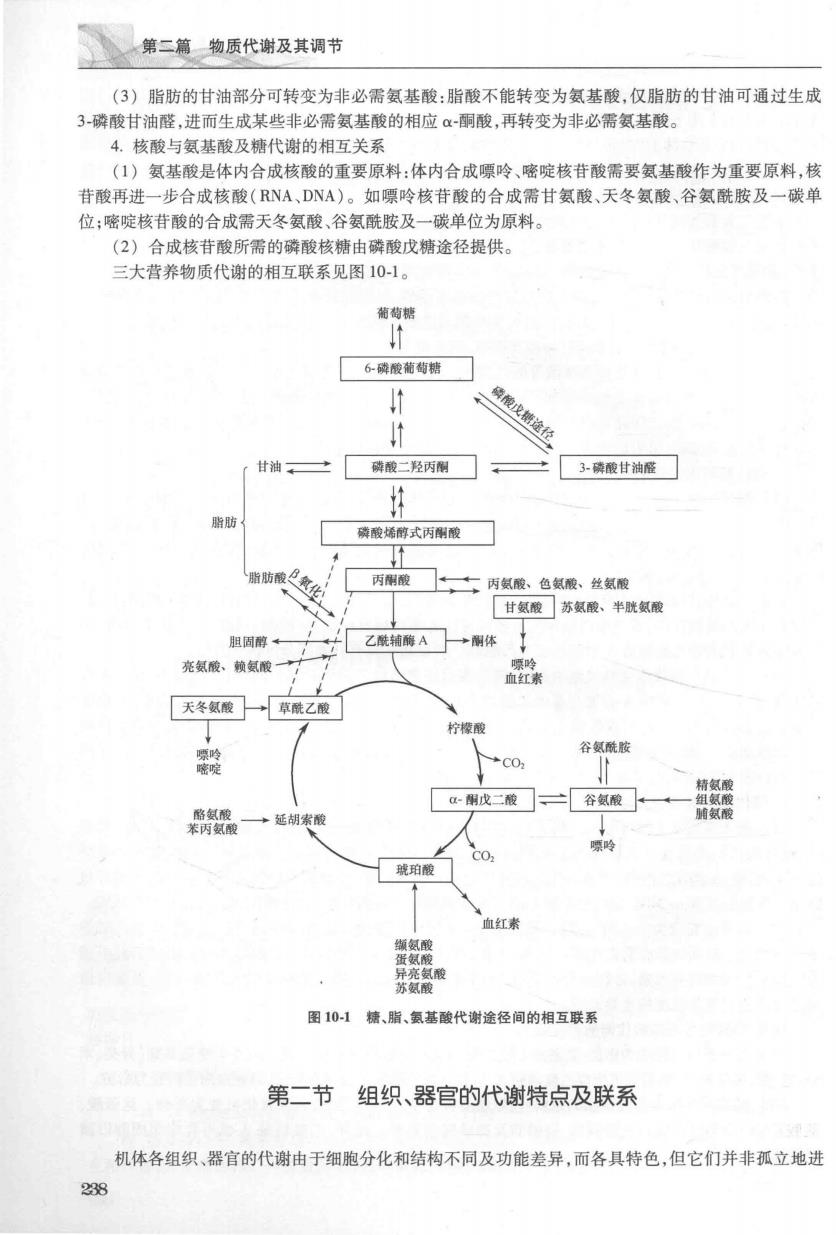
第二篇物质代谢及其调节 (3)脂肪的甘油部分可转变为非必需氨基酸:脂酸不能转变为氢基酸,仅脂肪的甘油可通过生成 3-磷酸甘油醛,进而生成某些非必需氨基酸的相应α-酮酸,再转变为非必需氨基酸。 4.核酸与氨基酸及糖代谢的相互关系 (1)氨基酸是体内合成核酸的重要原料:体内合成嘌吟、嘧啶核苷酸需要氨基酸作为重要原料,核 苷酸再进一步合成核酸(RNA、DNA)。如骠玲核苷酸的合成需甘氨酸、天冬氨酸、谷氨酰胺及一碳单 位;嘧啶核苷酸的合成需天冬氨酸、谷氨酰胺及一碳单位为原料。 (2)合成核苷酸所需的磷酸核糖由磷酸戊糖途径提供。 三大营养物质代谢的相互联系见图10-1。 葡萄糖 6砖棱葡萄糖 途程 甘油。二 磷酸二羟丙 3磷酸甘油 脂肪 磷酸烯醇式丙刚酸 丙酮酸 ☐上士丙氨酸、色氨酸、丝氨酸 甘氨酸苏氨酸、半胱氨酸 乙酰轴酶A一→体 充氨酸、赖氨酸 天冬氨酸→草酰乙酸 谷氨酰 C02 酮戊二酸]、三谷氨酸 玻珀酸 血红素 图101糖、脂、氨基酸代谢途径间的相互联系 第二节组织、器官的代谢特点及联系 机体各组织、器官的代谢由于细胞分化和结构不同及功能差异,而各具特色,但它们并非孤立地进 23

第十章物质代谢的联系与调节 行,而是通过血液循环及神经系统联成统一整体(表10-1)。 表101重要器官及组织氧化供能的特点 器官组织 特有的酶 功能 主要代谢途径 主要代谢物 主要代谢产物 葡萄糖激酶,葡萄糖-6磷代谢枢纽糖异生,脂酸B-氧葡萄糖,脂酸,乳酸,葡萄糖,VLDL, 酸酶,甘油激酶,磷酸烯 化,糖有氧氧化,酮甘油,氨基酸等 HDL,酮体 醇式丙酮酸羧激酶 体生成 神经中枢糖有氧氧化,氨基酸葡萄糖,氨基酸,C02,H20 代谢 酮体 肌肉 脂蛋白脂酶,呼吸链丰富收缩 糖酵解,有氧氧化 脂酸,葡萄糖,酮体乳酸,C02,H20 肾 甘油激酶,磷酸烯醇式丙排泄尿液糖异生,糖酵解,酮脂酸,葡萄糖,乳酸,葡萄糖 酮酸羧激酶 体生成 甘油 心 脂蛋白脂酶,呼吸链丰富泵出血液有氧氧化 乳酸,葡萄糖,VLDL C02,H20 脂肪组织脂蛋白脂酶,激素敏感脂储存及动酯化脂酸,脂解 VLDL,CM 游离脂酸,甘油 肪酶一 员脂肪 红细胞无线粒体 运输氧糖酵解 葡萄糖 乳酸 一、肝 肝是机体物质代谢的枢纽,是人体的“中心生化工厂”。它的耗0,量占全身耗0,量的20%。它不 仅在糖、脂、蛋白质、水、盐及维生素代谢中均具有独特而重要的作用,而且还有监控和调节血液化学组 成的功能。 (一)肝在糖代谢中的作用 1.肝内生成的葡糖6-磷酸是糖原合成及其他单糖转换的枢纽肝细胞含有葡萄糖转运蛋白2 (glucose transporter2,GLUT2),使肝细胞能有效摄取血中的葡萄糖,维持肝细胞与血糖几乎相等的浓 度。葡萄糖激酶仅在肝细胞内且其Km值比肝外组织己糖激酶的Km值高,这一特性使肝在高浓度葡 萄糖情况下将葡萄糖转化为葡糖6-磷酸,进而在合成糖原;而在葡萄糖浓度较低时却能抑制糖原合成, 以确保脑等重要组织糖供给。血中葡萄糖浓度过低时,肝糖原合成抑制而分解加强,补充血糖,供肝外 组织利用。故肝糖原合成/分解是血糖调节的重要机制之一。肝受损时,糖原转换能力降低,所以严重 肝病患者糖耐量能力下降,易出现餐后高血糖、空腹低血糖等症候。 此外,肝内生成的葡糖-6磷酸还是葡萄糖、果糖、半乳糖及甘露糖互变的枢纽物质。通过这一枢 纽,小肠吸收的其他单糖均可在肝内转变为葡萄糖进一步代谢。 2.肝是糖异生的主要场所肝糖原分解虽可补充血糖,但肝糖原储存有限(占肝重的10%,约 150g),故肝糖原的分解仅能持续16~24小时。较长时间不进食,肝糖异生则成为血糖的重要来源。肝 是糖异生的主要器官,可使氨基酸、乳酸、甘油等非糖物质转变为糖,以保证机体对糖的需要。即使在正 常情况下,每日经糖异生途径转变而来的葡萄糖仍可达80~160g。 (二)肝在脂代谢中的作用 1,肝是内源性甘油三酯合成的主要场所因肝细胞内的葡萄糖激酶K值很大,故肝细胞内的糖 酵解途径主要依赖葡萄糖的浓度,只有葡萄糖浓度较高时才会有部分葡萄糖进入酵解途径被肝利用。 通常情况下,肝能量供应以脂酸氧化为主。脂酸氧化而来的乙酰辅酶A除进入三羧酸循环氧化分解供 能,还可以作为酮体合成的原料。此外,葡萄糖供应过剩时,除少部分合成糖原,肝还可以将大量葡萄糖 来源的乙酰辅酶A转变成脂酸,进而合成甘油三酯,这是内源性甘油三酯的主要来源,并以VLDL形式 239