
第二十四章干细胞 学习目标 通过本章的学习,你应该能够: 掌握干细胞的概念和基本特性。 熟悉干细胞的分类及其特点。 了解干细胞临床应用及与医学的关系。 干细胞(stem cl)即为起源细胞,它是一类具有多向分化潜能和自我复制能力的原始的未分化细 胞,是在生物个体发育过程中存在于各种组织中的未成熟的或未分化的原始细胞。干细胞是调节个体 的生长发有,组织器官结构和功能的动态平衡,以及组织器官损伤后的修复再生等生命现象的细胞基 础。干细胞可分为胚胎干细胞和成体干细胞。 第一节干细胞的基本概念 干细胞是一类原始且未特化的多潜能细胞,具有自我复制的能力。在一定条件下,它可以分化成多 种功能细胞。在个体发育中,人类或哺乳动物所有的组织或器官中都有干细胞存在。虽然存在于不同 组织器官中的干细胞其生物学特性差异较大,但这些干细胞都具有自我更新的能力和分化产生特定功 能细胞的潜能,而且也都承担着调节相应组织器官结构和功能的动态平衡的作用。因此,认识它们的基 本特征,有助于了解各种干细胞在所在组织中的功能行为及其调控机制。 “stem c©”一词,早在1896年就首次出现在美国生物学家Wilson的一篇关于蠕虫发有的研究论文 中,用来描述存在于寄生虫生殖系的祖细胞(具有产生子代细胞的能力)。然而,直到最近的半个多世 纪,干细胞研究才得以迅猛发展。1998年,美国威斯康星大学Thomson等分离人的内细胞团细胞并成 功建立了人的胚胎干细胞系。与此同时,美国约翰·霍普金斯大学Gearhart等从人的原始生殖细胞中 建立了胚胎生殖细胞系。随后,以色列、澳大利亚、日本、新加坡等国家的科学家先后从体外受精卵建立 了人胚胎干细胞系,并诱导胚胎干细胞分化为神经细胞、造血细胞、肌肉细胞、胰岛细胞等,使得胚胎干 细胞的研究更加令人关注,并带动了世界范围内的干细胞研究热潮。这些研究也让人们看到干细胞生 物工程的曙光:可以在体外培育所需的细胞、组织甚至是器官,用来修复患者体内损伤的组织器官。 2006年,日本科学家Shinya Yamanaka通过逆转录病毒携带四种转录因子0ct3/4、Sox2、KI件和 c-Myc,将成纤维细胞重编程为胚胎干细胞样细胞,即诱导多能干细胞(induced pluripotent stem cell,iPS cell)。2007年,Yamanaka和美国科学家Thomson几乎同时报道,利用Ps技术同样可以诱导人皮肤成 纤维细胞去分化为几乎与人胚胎干细胞完全一样的多能干细胞。所不同的是Yamanaka依然采用逆转 录病毒引入Oct4,Sox2、c-Mye和K4四种因子组合,而Thomson采用慢病毒载体引人0ct4,Sox2、Nanog 534

第二十四章干细胞人 和Lin28四种因子组合。这两项研究成果,分别被Science和Nature杂志列为2007年十大科技进展中的 第一位和第二位。日本科学家Yamanaka也因在细胞重编程研究领域的开创性贡献,获得2012年诺贝 尔生理学或医学奖。2008年,哈佛大学的Dal©y利用PS技术,将来源于10种不同遗传病患者的皮肤 细胞重编程为S细胞。后续的研究进一步证实,这些建立的S细胞具有向内、中、外三个胚层分化的 能力,并可以进一步分化为特定的细胞系,这些S细胞将在疾病模型、药物筛选及细胞治疗等研究方 面发挥重要作用。 一、千细胞的分类 根据干细胞所处的发育阶段分为胚胎干细胞(embryonic stem cell,ES细胞)和成体干细胞(somatic stem cell)。根据干细胞的发育潜能分为三类:全能干细胞(totipotent stem cell,TSC)、多能干细胞(pluri- potent stem cel)和单能干细胞(unipoter nce)(专能干细胞)。干细胞(stem cell)是一种未充分分 化,尚不成熟的细胞,具有再生各种组织器官和人体的潜在功能,医学界称为“万用细胞”。全能干细 胞:具有能够发育成为各种组织器官的完整个体潜能的细胞。全能干细胞是指受精卵到卵裂期8细胞 前的所有细胞。胚胎干细胞在进一步的分化中,可形成各种组织干细胞,又称多能干细胞,它具有分化 出多种细胞组织的潜能,但不能发育成完整的个体。多能干细胞取自囊胚,原肠胚期。多能干细胞进 步分化,可形成专能干细胞,专能干细胞只能分化成某一类型的细胞。原肠胚以后的干细胞只能是专能 干细胞了,如某些肝脏细胞,骨随造血千细胞。所以脐带或者成人骨髓中的都已经是专能干细胞了,即 纯体外培养只能分裂分化出特定的组织细胞,如骨髓只能分裂出各种血细胞。 胚胎干细胞(embryonic stem cell,S细胞)是一种高度未分化细胞,早期胚胎(原肠胚期之前)或原 始性腺中分离出来的一类细胞,它具有体外培养无限增殖、自我更新和多向分化的特性。无论在体外还 是体内环境,S细胞都能被诱导分化为机体几乎所有的细胞类型,包括生殖细胞 成体干细胞(somatic stem cell)是指存在于一种已经分化组织中的未分化细胞.这种细胞能够自我 更新并且能够特化形成组成该类型组织的细胞。然而,这个观点目前受到了挑战,最新的研究表明,组 织特异性干细胞同样具有分化成其他细胞或组织的潜能,这为干细胞的应用开创了更广泛的空间。成 体干细胞存在于机体的各种组织器官中。成年个体组织中的成体干细胞在正常情祝下大多处于休眠状 态,在病理状态或在外因诱导下可以表现出不同程度的再生和更新能力。成年动物的许多组织和器官 比如表皮和造血系统,具有修复和再生的能力。成体干细胞在其中起着关键的作用。在特定条件下,成 体干细胞或者产生新的干细胞,或者按一定的程序分化,形成新的功能细胞,从而使组织和器官保持生 长和衰退的动态平衡。 诱导多能干细胞(induced pluripotent stem cells,.iPS cells),是指通过基因转染技术将某些转录因子 导人动物或人的体细胞,使体细胞直接重构成为胚胎干细胞细胞样的多潜能细胞。 二、干细胞的形态、生化和增殖,分化特征 (一】干细胞的形态特征 干细胞在形态上具有共性,通常呈圆形或椭圆形,细胞体积小,核质比例相对较大,核仁明显,细服 核多为常染色质,细胞质中各种细胞器(如内质网、高尔基复合体及线粒体等)不发达等等。 (二)干细胞的生化特征 在目前的研究中,干细胞的生化特性常被作为鉴定干细胞在组织中存在和评价其分化程度的标志 干细胞都具有较高的端粒酶活性,不同的干细胞具有不同的生化标志。例如,nsin为神经干细胞的标 志分子;角蛋白15是确定毛囊中表皮干细胞的标志分子:CD34是造血干细胞的标志分子;人胚胎干细 胞表达阶段特异性胚胎抗原((stage-specific embryonic antigens,SSEA-3和SSEA-4)及Nanog,具有碱性 酸酶活性等。但是,不能仅仅根据细胞的形态和表面抗原来鉴定干细胞,具有增殖和自我更新能力以及 在适当条件下表现出一定的分化潜能才是干细胞的本质特征。干细胞的生化特性还与其分化程度有 535

第五篇细胞分裂增殖与生长发育 关。例如,造血干细胞的端粒酶(telomerase)活性很高(可达到造血系统肿瘤细胞的端粒酶活性水平) 但当它分化为前体细胞后,其瑞粒酶活性便随之降低。 (三)干细胞的增殖特性 1.干细胞增殖的缓慢性 当干细胞进入分化程序后,首先经过一个短暂的增殖期,产生过渡放大 细胞(transit amplifying cell)。过渡放大细胞经若干次分裂后,便可产生分化细胞。过渡放大细胞的生 物学意义在于它可以减少干细胞的分裂次数,而产生较多的分化细胞。细胞动力学研究表明,干细胞本 身的增殖速率通常是很慢的,而组织中的那些过渡细胞的分裂速度则相对较快。如小肠干细胞较其过 渡细胞的分裂速度大约慢一倍。目前认为,这种缓慢增殖的特性有利于干细胞对特定的外界信号做出 反应,以决定细胞是进入增殖状态或是进入特定的分化程序。这种缓慢增殖的特性还可以减少基因突 变的危险,使干细胞有更多的时间发现和校正复制错误。 2.干细胞增殖的自稳定性 自稳定性(self-maintenance)是指干细胞具有自我更新(self-renewing 并维特其自身数目恒定的特性,它是干细胞的基本特征之一。当干细胞发生分裂后,如果所产生的两个 子代细胞都是干细胞或都是分化细胞,这种分裂方式称为对称分裂(y v击rision):加果产生一个 干细胞和一个分化细胞,则称之为不对称分裂(asymmetry division)。对无脊椎动物而言,不对称分裂是 干细胞维持自身数目恒定的基本方式,但对哺乳动物而言,却并非如此简单,在大多数哺乳动物的可自 我更新的组织中,干细胞分裂产生的两个子细胞既可能是两个干细胞,也可能是两个特定分化细胞,因 此哺乳动物的单个干细胞通常是讲行对称分裂。但平均而言,每一个干细胞可以产生一个子代干细胞 和一个特定分化细胞。因此,哺乳动物的干细胞是种群(而不是单个干细胞)意义上的不对称分裂,称 之为种群不对称分裂(populational asymmet女y division)(图24-l)。 )多能干细胞 干细 前体细 自我更新 分化 分化的 图241干细胞的不对称分裂和分化示意图 (四)干细胞的分化特性 1.干细胞具有分化为一种以上特定功能细胞的潜能具有分化为特定功能细胞的能力是干细 的另一个本质特征。如上所述,处于不同发育阶段的干细胞其分化潜能各不相同。例如,受精卵是全能 干细胞,具有形成一个完整个体的能力:囊胚中的内细胞团细胞是多能干细胞,能产生个体中任何一种 组织细胞的潜能,但不能产生胎盘和其他一些发育时所必需的支持组织:存在于胚胎各组织器官原基中 的干细胞或成熟个体组织器官中的成体干细胞属于单能干细胞,其分化潜能进一步受到限制,通常只能 分化为参与其相应组织器官组成的细胞。 2.干细胞的分化具有一定的可塑性曾经认为成体干细胞只能向一种类型或与之密切相关的细 胞分化。然而,越来越多的证据已经表明,成体干细胞具有一定的可塑性,在适当的条件下可以表现出 536
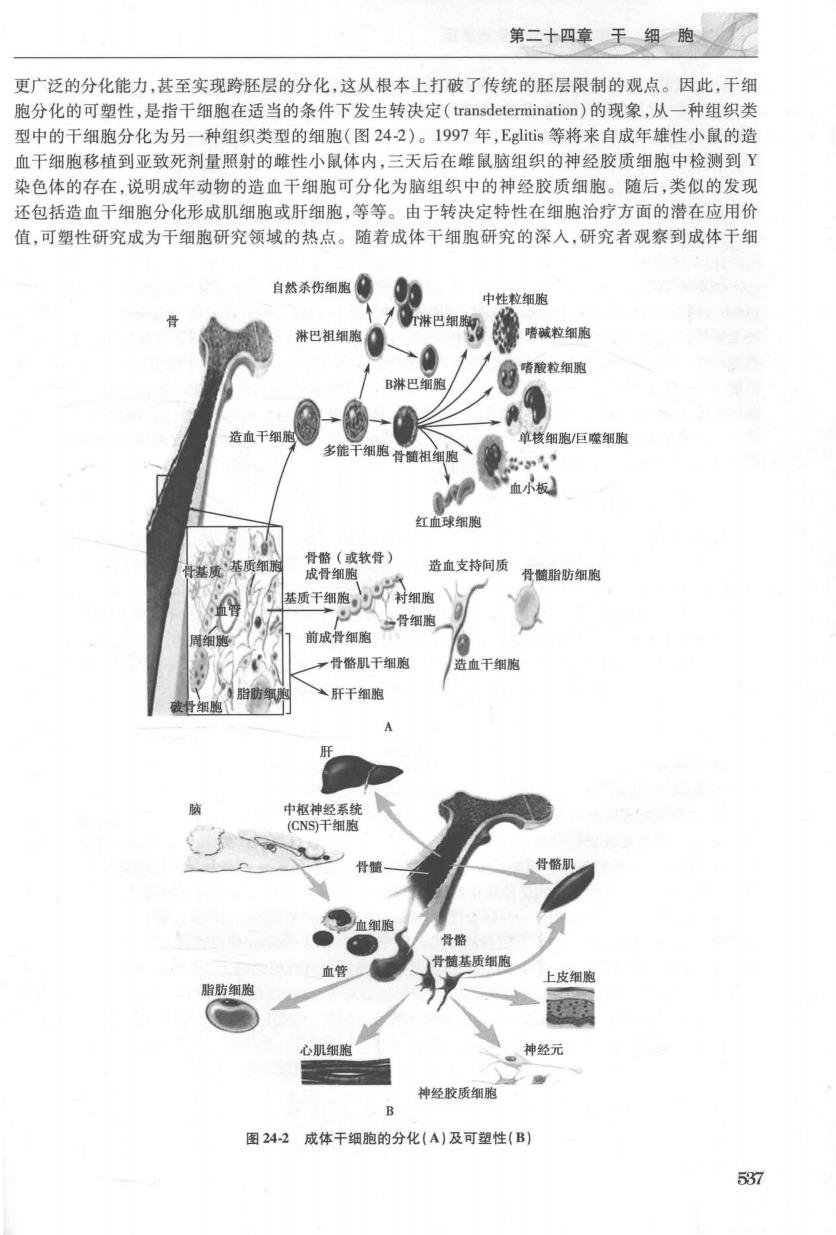
第二十四章于绵渔 更广泛的分化能力,甚至实现跨胚层的分化,这从根本上打破了传统的胚层限制的观点。因此,干细 胞分化的可塑性,是指干细胞在适当的条件下发生转决定(transdetermination)的现象,从一种组织类 型中的干细胞分化为另一种组织类型的细胞(图24-2)。1997年,Egs等将来自成年雄性小鼠的造 血干细胞移植到亚致死剂量照射的雌性小鼠体内,三天后在雕鼠脑组织的神经胶质细胞中检测到Y 染色体的存在,说明成年动物的造血干细胞可分化为脑组织中的神经胶质细胞。随后,类似的发现 还包括造血干细胞分化形成肌细胞或肝细胞,等等。由于转决定特性在细胞治疗方面的潜在应用价 值,可塑性研究成为干细胞研究领域的热点。随着成体干细胞研究的深人,研究者观察到成体干细 自然杀伤细胞● 的防细 心肌细 。神经元 神经胶质细胞 图242成体干细胞的分化(A)及可塑性(B) 537

】第五分与生长发有 胞可以突破其“发育限制性”,跨系,甚至跨胚层分化为其他类型组织细胞。例如,骨髓来源的干细胞 在特定环境中可向肝脏、胰腺、肌肉及神经细胞分化:肌肉、神经干细胞也可向造血细胞分化。人们 称这种现象为“干细胞的可塑性”。成体干细胞的这种特性为其在多种难治性、终末期疾病的治疗中 应用带来希望。 第二节胚胎干细胞 胚胎干细胞(embryonie stem cell,ES细胞)。胚胎干细胞当受精卵分裂发育成囊胚时,内层细胞团 (inner cell mass)的细胞即为胚胎干细胞。胚胎干细胞(ES细胞)是一种高度未分化细胞。它具有发有 的全能性,能分化出成体动物的所有组织和器官,包括生殖细胞。研究和利用S细胞是当前生物工程 领域的核心问题之 一。ES细胞的研究可追湖到20世纪50年代,由于畸胎瘤干细胞(EC细胞)的发现 开始了ES细胞的生物学研究历程。1981年,Evas首次分离并成功培养小鼠ES细胞,奠定了胚胎干细 胞体外研究的技术基础。随后,陆续有猪、牛,绵羊、仓鼠、鸡、斑马鱼、恒河猴等脊椎动物的胚胎干细胞 得以成功分离和培养。1998年,Thomson从体外受精获得的人囊胚内细胞团中,分离培养人胚胎干细胞 获得成功,为胚胎干细胞走向临床应用奠定了基础。 一、胚胎干细胞简介 胚胎干细胞通常是指存在于囊胚期胚胎中的内细胞团细胞。在囊胚后续的发育过程中,有些类型 的细胞在特定的条件下也可表现出类似于ES细胞的特性,即自我更新能力和多能性分化潜能等。1998 年,美国约翰·蛋普金斯大学Gearhart从5~9周龄的流产胎儿的生殖蝽中分离培养了这种细胞,并在 体外证明了它与来自于内细胞团的S细胞的具有相似的生物学特性。另外,小鼠的卵黄囊细胞在一定 的诱导条件下也可以表现出类似地多向分化潜能。胚胎干细胞的来源一般有5方面:①囊胚的内细胞 团细胞;②胚胎生殖嵴处的胚胎生殖细胞:③终止妊娠的胎儿组织中分离出多能干细胞:④在体外用体 细胞核转移技术获得胚胎干细胞;⑤从恶性胚胎肿瘤或畸胎瘤细胞获得。 二、胚胎干细胞的特征 S细胞具有与早期胚胎细胞相似的形态结构,细胞核大,有一个或几个核仁,胞核中多为常染色 质,胞质胞浆少,结构简单。体外培养时,细胞排列紧密,呈集落状生长。用碱性磷酸酶染色,S细胞 棕红色,而周围的成纤维细胞呈谈黄色。细胞克隆和周围存在明显界限,形成的克隆细胞彼此界限不 清,细胞表面右折光较强的脂状小滴。细胞克隆形态多样,多数呈岛状或巢状。ES细胞的全能性指S 细胞在解除分化抑制的条件下能参与包括生殖腺在内的各种组织的发育潜力,即S细胞具有发有成完 整动物体的能力,可以为细胞的遗传操作和细胞分化研究提供丰富的试验材料。ES细胞发育全能性的 标志是ES细胞表面表达时相专一性胚胎抗原(stage specific embryonicant,SSEA),而且可以检查到Oc4 基因的表达,这两种蛋白是发育全能性的标志。ES细胞中AKP及端粒酶活性较高,可用于ES细胞分 化与否的鉴定。S细胞的多能性是指S细胞具有发育成多种组织的能力,参与部分组织的形成。将 S细胞培养在不含分化抑制物的培养基上,可以形成类胚体。将S细胞在特定培养基进行培养,可以 定向分化成特定组织,如ES细胞在含有白血病抑制因子(LF)和维生素A酸(RA)的培养基上,可以分 化形成全壁内胚层,将ES细胞与胚胎细胞共培养或将ES细胞注入囊胚腔中,ES细胞就会参与多种组 织的发育。 三、胚胎干细胞具有向三个胚层组织细胞分化的潜能 由于胚胎干细胞体外培养的成功,这一概念已经通过实验得到了充分的证实,这些实验一般包括三