
第12章蛋白质的生物合成 学习要求 续表 能量 ATP ,GTP 掌握翻译的概念,蛋白质生物合成体系的组成及 无机离子 作用。 Me2、K 掌握遗传密码的概念,特点、起始密码子与终止 1.mRNA是蛋白质生物合成的直接模板 密码子。热悉核糖体在蛋白质生物合成中的作用,热 悉核糖体上AP,E位及其作用,了解核糖体的组成。 (1)mRNA的结构特点:mRNA可以区分为两端 的非翻译区和中间的编码区(开放阅读框),参见 掌握tRNA在蛋白质生物合成中的作用,熟悉tRNA 下图。 与蛋白质生物合成右关的关键部位(氨其酸接受赠 从mRNA5'.的起始密码子到3'的终止密码子之 反密码子) 专肽,转位酶及其作用,了解蛋日 质生物合成中蛋白质因 间的核苷酸序列,称为开放阅读框(open reading 能源物质及其作用。掌掘 fame,ORF)。 氨基酸活化的概念及氨基酰RNA合成酶的作用特 点。了解氨基酰RNA的表示方法与起始氨基酰-R mRNA 5.区 编码区或ORF NA 熟悉原核生物SD序列进位、成肽转位的概 Translation 了解原核生物多肽链合成的过程,了解各阶段蛋 白质因子的作用、能量消耗情 了解真核生物多 protein N ■C流 链合成 过程与原核生物的主要差别。了解多聚核糖 真核生物和原核生物的mRNA的结构明显不同, 体的概念。 参见表12.2。 了解蛋白质生物合成后的加工修饰方式。了解 表12-2原核生物与真核生物mRNA的比较 蛋白质靶向输送至细胞特定部位的方式。熟悉信号 原核生物 肽的概念与作用 真核生物 了解于扰蛋白质生物合成的抗生素及其作用,了 稳定性 不稳定,半衰期2秒较稳定,半衰期几个小 解白喉毒素、干扰素作用的机理。 到几分钟 时到24小时 讲义要点 合成部位 胞液中合成,不需核质中合成,需转移到 转移 细胞质中 (一)蛋白质生物合成概述 结构 5'端无“帽子”结构 5'端有“帽子”结构,3 蛋白质的生物合成也称翻译(transation),是以 3”端无或者只不 端有py(A)尾巴 mRNA为模板,把mRNA上的核苷酸序列解读为蛋白 较短的p(A) 结构 质的氨基酸序列.从而合成蛋白质的过程。 编码蛋白质同时编码几种不同的只编码一种蛋白质:单 mRNA作为直接的模板,但真正的遗传信息或指 蛋白质:多顺反子 顺反子 今是心在在DNA序刚中 (二)蛋白质生物合成体系(表12-1) 翻译起始相SD序列 关列 表121蛋白质生物合成体系 模板 mRNA (2)遗传密码 1)概念:mRNA分子上从5'至3'方向,每三个相 原料及运输工具 20种氨基酸,tRNA 部的核苷酸为一组,在蛋白质生物合成中代表某种氨 合成场所 核糖体(RNA+蛋白质) 及蛋白质因子 氨基酰RA合成酶,肽酰基转移 基酸或肽链合成的起始或终止信号,称为遗传密码。 从另一种角度理解,遗传密码就是一种规则,它 起始/延长放因子等 规定了mRNA中的核苷酸序列如何解读成蛋白质的 ·126:
126 第 12 章 蛋白质的生物合成 学 习 要 求 掌握翻译的概念、蛋白质生物合成体系的组成及 作用ꎮ 掌握遗传密码的概念、特点、起始密码子与终止 密码子ꎮ 熟悉核糖体在蛋白质生物合成中的作用ꎬ熟 悉核糖体上 A、P、E 位及其作用ꎬ了解核糖体的组成ꎮ 掌握 tRNA 在蛋白质生物合成中的作用ꎬ熟悉 tRNA 与蛋白质生物合成有关的关键部位(氨基酸接受臂、 反密码子)ꎮ 熟悉转肽酶、转位酶及其作用ꎬ了解蛋白 质生物合成中蛋白质因子、能源物质及其作用ꎮ 掌握 氨基酸活化的概念及氨基酰 ̄tRNA 合成酶的作用特 点ꎮ 了解氨基酰 ̄tRNA 的表示方法与起始氨基酰 ̄tR ̄ NAꎮ 熟悉原核生物 S ̄D 序列、进位、成肽、转位的概 念ꎬ了解原核生物多肽链合成的过程ꎬ了解各阶段蛋 白质因子的作用、能量消耗情况ꎮ 了解真核生物多肽 链合成过程与原核生物的主要差别ꎮ 了解多聚核糖 体的概念ꎮ 了解蛋白质生物合成后的加工修饰方式ꎮ 了解 蛋白质靶向输送至细胞特定部位的方式ꎮ 熟悉信号 肽的概念与作用ꎮ 了解干扰蛋白质生物合成的抗生素及其作用ꎬ了 解白喉毒素、干扰素作用的机理ꎮ 讲 义 要 点 (一) 蛋白质生物合成概述 蛋白质的生物合成也称翻译( translation)ꎬ是以 mRNA 为模板ꎬ把 mRNA 上的核苷酸序列解读为蛋白 质的氨基酸序列ꎬ从而合成蛋白质的过程ꎮ mRNA 作为直接的模板ꎬ但真正的遗传信息或指 令是贮存在 DNA 序列中ꎮ (二) 蛋白质生物合成体系(表 12 ̄ 1) 表 12 ̄ 1 蛋白质生物合成体系 模板 mRNA 原料及运输工具 20 种氨基酸ꎬtRNA 合成场所 核糖体(rRNA+蛋白质) 酶及蛋白质因子 氨基酰 tRNA 合成酶ꎬ肽酰基转移 酶ꎬ起始/ 延长/ 释放因子等 续表 能量 ATP、GTP 无机离子 Mg 2+ 、 K + 1 mRNA 是蛋白质生物合成的直接模板 (1) mRNA 的结构特点:mRNA 可以区分为两端 的非翻译区和中间的编码区 ( 开放阅读框)ꎬ参见 下图ꎮ 从 mRNA 5′ ̄的起始密码子到 3′ ̄的终止密码子之 间的 核 苷 酸 序 列ꎬ 称 为 开 放 阅 读 框 ( open reading frameꎬORF)ꎮ 真核生物和原核生物的 mRNA 的结构明显不同ꎬ 参见表 12 ̄ 2ꎮ 表 12 ̄ 2 原核生物与真核生物 mRNA 的比较 原核生物 真核生物 稳定性 不稳定ꎬ半衰期 2 秒 到几分钟 较稳定ꎬ半衰期几个小 时到 24 小时 合成部位 胞 液 中 合 成ꎬ 不 需 转移 核质中合成ꎬ需转移到 细胞质中 结构 5’端无“帽子”结构ꎬ 3’ 端 无 或 者 只 有 较 短 的 poly ( A ) 结构 5’端有“帽子”结构ꎬ3’ 端有 poly(A)尾巴 编码蛋白质 同时编码几种不同的 蛋白质ꎻ多顺反子 只编码一种蛋白质ꎻ单 顺反子 翻译起始相 关序列 SD 序列 “帽子” 结构及起始密 码 AUG (2) 遗传密码 1) 概念:mRNA 分子上从 5′至 3′方向ꎬ每三个相 邻的核苷酸为一组ꎬ在蛋白质生物合成中代表某种氨 基酸或肽链合成的起始或终止信号ꎬ称为遗传密码ꎮ 从另一种角度理解ꎬ遗传密码就是一种规则ꎬ它 规定了 mRNA 中的核苷酸序列如何解读成蛋白质的
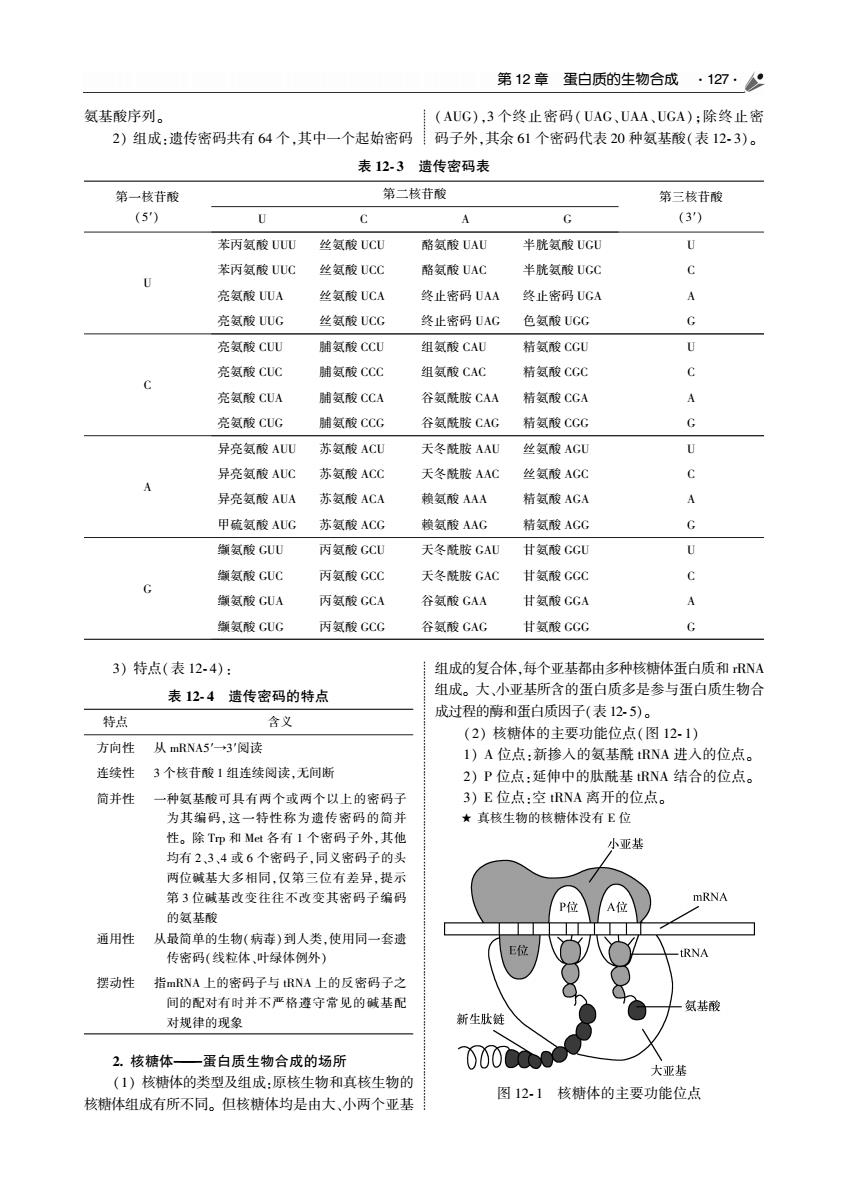
第12章蛋白质的生物合成·127·% 氨基酸序列。 (AUG),3个终止密码(UAG、UAA,UGA):除终止密 2)组成:遗传密码共有64个,其中一个起始密码码子外,其余61个密码代表20种氨基酸(表12-3)。 表12.3遗传密码表 第一苷酸 第二枝苷酸 第三核苷酸 (5) U A (3) 苯丙氨酸UUWU 丝氨酸UCU 格氨酸UAU 半酰氨酸UCL 苯丙氨酸UUC 丝氨酸UCC 酪氨酸UAC 半胱氢酸UGC 亮红酸UUA 丝氨酸UC 终止密码UAA 终止密码UGA 亮氨酸UUG 丝氨酸UCC 终止密码UAC 色氢酸CC 亮氨酸CUU 铺酸CCLU 组氨酸CAU 精氨酸CCU 亮氨酸CLUC 脯氨酸CCC 组氨酸CAC 精氨酸CGC 亮氢酸CUA 脯氢酸CCA 谷氨酰胺CAA 精氢酸CGA A 亮氨酸CUG 脯氨酸CCC 谷氨酰胺CAG 精氨酸CCG 异亮氨酸AUU 苏氨酸AC 天冬酰胺AAU 丝氨酸AGU U 异亮氨酸ALC 苏氨酸ACC 天冬酰胺AAC 丝氨酸ACC 异亮氨酸AUA 苏复酸ACA 轮氨酸AAA 精氨酸AGA A 甲硫氨酸AUG 苏氨酸ACG 氨酸AAG 精氨酸AGC G 挥氨酸CUU 丙氨酸CC 天察胜安GAU 甘氨酸CC可 丙氨酸GCC 天冬酰胺GAC 甘氨酸GCC 徽氨酸GUA 丙氨酸GCA 谷氨酸CGAA 甘氨酸CCA A 犧氨酸GUG 丙氨酸GCG 谷氨酸GAG 甘氨酸GGG G 3)特点(表12-4) 组成的复合体,每个亚基都由多种核糖体蛋白质和RNA 表12-4造传密码的特点 组成。大,小亚基所含的蛋白质多是参与蛋白质生物合 成过程的南和蛋白质因子(表12.5)。 特点 含义 (2)核糖体的主要功能位点(图12.1) 方向性从mRNA5'→3阅读 入的位点 连续性 3个核苷酸1组连续阅读,无间断 A位点:新擦人的氢装酰RN 2)P位点:延伸中的肽酰基RNA结合的位点。 简并性 一种氨基酸可具有两个或两个以上的密码子 3)E位点:室RNA离开的位点。 为其编码,这一特性称为遗传密码的简 ★真核生物的核糖体没有E位 性。除Tp和Mt各有I个密码子外,其他 小亚基 均有2,3、4或6个密码子,问义密码子的头 两位碱基大多相同,仅第三位有差异,提万 第3位碱基改变往往不改变其密码子编码 A位 的虱基腹 通用性 从最简单的生物(病毒)到人类,使用同一套遗 传密码(线粒体,叶绿体例外) 摆动性指mRNA上的密码子与RNA上的反密码子之 间的配对有时并不严格蓬守常见的碱基配 对规律的现象 新生金 2.核糖体- 一蛋白质生物合成的场所 000Cn 大亚基 (1)核糖体的类型及组成:原核生物和真核生物的 核糖体组成有所不同。但核糖体均是由大、小两个亚精 图121核糖体的主要功能位点
第 12 章 蛋白质的生物合成 127 氨基酸序列ꎮ 2) 组成:遗传密码共有 64 个ꎬ其中一个起始密码 (AUG)ꎬ3 个终止密码(UAG、UAA、UGA)ꎻ除终止密 码子外ꎬ其余 61 个密码代表 20 种氨基酸(表 12 ̄ 3)ꎮ 表 12 ̄ 3 遗传密码表 第一核苷酸 (5′) 第二核苷酸 U C A G 第三核苷酸 (3′) U 苯丙氨酸 UUU 丝氨酸 UCU 酪氨酸 UAU 半胱氨酸 UGU U 苯丙氨酸 UUC 丝氨酸 UCC 酪氨酸 UAC 半胱氨酸 UGC C 亮氨酸 UUA 丝氨酸 UCA 终止密码 UAA 终止密码 UGA A 亮氨酸 UUG 丝氨酸 UCG 终止密码 UAG 色氨酸 UGG G C 亮氨酸 CUU 脯氨酸 CCU 组氨酸 CAU 精氨酸 CGU U 亮氨酸 CUC 脯氨酸 CCC 组氨酸 CAC 精氨酸 CGC C 亮氨酸 CUA 脯氨酸 CCA 谷氨酰胺 CAA 精氨酸 CGA A 亮氨酸 CUG 脯氨酸 CCG 谷氨酰胺 CAG 精氨酸 CGG G A 异亮氨酸 AUU 苏氨酸 ACU 天冬酰胺 AAU 丝氨酸 AGU U 异亮氨酸 AUC 苏氨酸 ACC 天冬酰胺 AAC 丝氨酸 AGC C 异亮氨酸 AUA 苏氨酸 ACA 赖氨酸 AAA 精氨酸 AGA A 甲硫氨酸 AUG 苏氨酸 ACG 赖氨酸 AAG 精氨酸 AGG G G 缬氨酸 GUU 丙氨酸 GCU 天冬酰胺 GAU 甘氨酸 GGU U 缬氨酸 GUC 丙氨酸 GCC 天冬酰胺 GAC 甘氨酸 GGC C 缬氨酸 GUA 丙氨酸 GCA 谷氨酸 GAA 甘氨酸 GGA A 缬氨酸 GUG 丙氨酸 GCG 谷氨酸 GAG 甘氨酸 GGG G 3) 特点(表 12 ̄ 4): 表 12 ̄ 4 遗传密码的特点 特点 含义 方向性 从 mRNA5′→3′阅读 连续性 3 个核苷酸 1 组连续阅读ꎬ无间断 简并性 一种氨基酸可具有两个或两个以上的密码子 为其编码ꎬ这一特性称为遗传密码的简并 性ꎮ 除 Trp 和 Met 各有 1 个密码子外ꎬ其他 均有 2、3、4 或 6 个密码子ꎬ同义密码子的头 两位碱基大多相同ꎬ仅第三位有差异ꎬ提示 第 3 位碱基改变往往不改变其密码子编码 的氨基酸 通用性 从最简单的生物(病毒)到人类ꎬ使用同一套遗 传密码(线粒体、叶绿体例外) 摆动性 指mRNA 上的密码子与 tRNA 上的反密码子之 间的配对有时并不严格遵守常见的碱基配 对规律的现象 2 核糖体———蛋白质生物合成的场所 (1) 核糖体的类型及组成:原核生物和真核生物的 核糖体组成有所不同ꎮ 但核糖体均是由大、小两个亚基 组成的复合体ꎬ每个亚基都由多种核糖体蛋白质和 rRNA 组成ꎮ 大、小亚基所含的蛋白质多是参与蛋白质生物合 成过程的酶和蛋白质因子(表 12 ̄ 5)ꎮ (2) 核糖体的主要功能位点(图 12 ̄ 1) 1) A 位点:新掺入的氨基酰 tRNA 进入的位点ꎮ 2) P 位点:延伸中的肽酰基 tRNA 结合的位点ꎮ 3) E 位点:空 tRNA 离开的位点ꎮ ★ 真核生物的核糖体没有 E 位 图 12 ̄ 1 核糖体的主要功能位点

·128·生物化学与分子生物学学习纲要与同步练习 表12.5原核生物与真核生物核糖体的组成 来源 沉降系数 重量(道尔倾) 亚基 RNA 含蛋白质种数 直技生物 77-809 3.6×10 18 33 5.5.85.28s 50 原核生物 70s 2.6×10 30S(小) 16S 21 50S(大) 235.5s 34 3.tRNA 氨基酸的运载工具及蛋白质生物合 1)标准配对:A=U.C=G 1#,2# 成的话配器 2)非标准配对:I一→AVC/U:U一→AVG等 3# (1)RNA是氨基酸的运载工具:胞液中的氨基 酸需要由RNA搬运到核糖体上才能组装成多肽链。 AA 1)氨基酸的活化 基酰RNA的合成 由氨基酰RNA合成酶催化,氨基酸的羧基与R NA3'未端CCA腺苷酸的3'0H以酯键连接,形成氨 3(② 基酰-RNA。 每个氨基酸活化需消耗1个ATP分子(2个高能 mRNA 5' TXYA☐ 磷酸键:ATP一AMP)。 氨基酸+RNA+AP氨基酸RNA合成氨琴 图12.2mRNA密码子与RNA反密码子的配对 酰-RNA+AMP+PP 表12-7密码子与反密码子配对的摆动现象 2)氨基酰-RNA合成酶的特性 RNA反密码子的第一位碱基 C A U G A高度专一性:氨基酰-RNA合成醇对底物氨基 酸和RNA都能高度特异性的识别,因此~密码 密码子的第三位碱基 G U AG CU UCA (mRNA)+反密码(RNA)+氨基酸”的结合是一种特 3.卷与蛋白质生物合成的和蛋白因子 异性结合 B具有校正活性:错误的氨基酸加人后会被水 (1)主要的醇 去掉 1)氢基酰RA合成酶:位于胞液中,催化氨基 上述两种机制保证了蛋白质合成的忠实性。 酸的活化。 2)转肽酶:是核糖体大亚基的成分,催化P位与 3)真核生物起始氨基酰-RNA是Met-RNA A位的氨基酸之间生成肽键 原核生物的起始密码只能辨认甲酰化的甲硫氨酸,即 N甲酰甲硫氨酸。 3)转位酶:其活性存在于延长因子G中,催化核 萌体的移位 表12-6原核生物与真核生物的起始氨基酸tRNA (2)蛋白质因子:见表128。 原核生物 真核生物 表128参与蛋白质生物合成的各种蛋白因子 起始甲酰甲硫氨酸(BM)、甲硫氨酸(Met)、MtR 原核生物 Met-RNA 真核生物 NA 起始因子 3种(F-1,F-2、10余种(至少9种 廷长Met-RNAM Met-RNA clF1-9) (2)tRNA是蛋白质生物合成的话配器:tRNA矩 延长因子 3粼EFRT、3种(EF1-a、eEFI EF-C) By、cEF.2) 借自身的反密码子与mRNA上的密码子通过碱基互 补配对作用相万识别使不同的氨甚酸按照密码子决 释因子 3种(R.1,R.21种(eRF) 定的次序合成多肽链,这也就是RNA的适配器作用 RF-3) 即mRNA序列中密码子的排列顺序通过RNA被转换 成多肽链中氨基酸的排列顺序。 (三)蛋白质的生物合成过程 由于配对具摆动性,因此一种RNA分子可识别 1.起始阶段 一种以上的同义密码子。 (1)参与起始的因子:见表12-9。 密码子与反密码子的配对方式(图12.2.表12.7)
128 生物化学与分子生物学学习纲要与同步练习 表 12 ̄ 5 原核生物与真核生物核糖体的组成 来源 沉降系数 重量(道尔顿) 亚基 rRNA 含蛋白质种数 真核生物 77~ 80S 3.6×10 6 40S(小) 60S(大) 18S 5Sꎬ 5.8Sꎬ 28S 33 50 原核生物 70S 2.6×10 6 30S(小) 50S(大) 16S 23Sꎬ 5S 21 34 3 tRNA———氨基酸的运载工具及蛋白质生物合 成的适配器 (1) tRNA 是氨基酸的运载工具:胞液中的氨基 酸需要由 tRNA 搬运到核糖体上才能组装成多肽链ꎮ 1) 氨基酸的活化———氨基酰 ̄tRNA 的合成ꎮ 由氨基酰 ̄tRNA 合成酶催化ꎬ氨基酸的羧基与 tR ̄ NA3′ ̄末端 ̄CCA 腺苷酸的 3′ ̄OH 以酯键连接ꎬ形成氨 基酰 ̄tRNAꎮ 每个氨基酸活化需消耗 1 个 ATP 分子(2 个高能 磷酸键:ATP→AMP)ꎮ 氨基酸+tRNA+ATP 氨基酸 ̄tRNA 合成酶 →氨基 酰 ̄tRNA+AMP+PPi 2) 氨基酰 ̄tRNA 合成酶的特性 A 高度专一性:氨基酰 ̄tRNA 合成酶对底物氨基 酸和 tRNA 都 能 高 度 特 异 性 的 识 别ꎬ 因 此 “ 密 码 (mRNA)+反密码( tRNA) +氨基酸” 的结合是一种特 异性结合ꎮ B 具有校正活性:错误的氨基酸加入后会被水解 去掉ꎮ 上述两种机制保证了蛋白质合成的忠实性ꎮ 3) 真核生物起始氨基酰 ̄tRNA 是 Met ̄tRNA Met i : 原核生物的起始密码只能辨认甲酰化的甲硫氨酸ꎬ即 N ̄甲酰甲硫氨酸ꎮ 表 12 ̄ 6 原核生物与真核生物的起始氨基酸 ̄tRNA 原核生物 真核生物 起始 甲酰甲硫氨酸( fMet)、 fMet ̄tRNA fMet 甲硫氨酸( Met)、Met ̄tR ̄ NA Met i 延长 Met ̄tRNA Met Met ̄tRNA Met e (2) tRNA 是蛋白质生物合成的适配器:tRNA 凭 借自身的反密码子与 mRNA 上的密码子通过碱基互 补配对作用相互识别ꎬ使不同的氨基酸按照密码子决 定的次序合成多肽链ꎬ这也就是 tRNA 的适配器作用ꎬ 即 mRNA 序列中密码子的排列顺序通过 tRNA 被转换 成多肽链中氨基酸的排列顺序ꎮ 由于配对具摆动性ꎬ因此一种 tRNA 分子可识别 一种以上的同义密码子ꎮ 密码子与反密码子的配对方式(图 12 ̄ 2ꎬ表 12 ̄7): 1) 标准配对: A=UꎬC≡G 1#ꎬ 2# 2) 非标准配对:I→A/ C/ Uꎻ U→A/ G 等 3# 图 12 ̄ 2 mRNA 密码子与 tRNA 反密码子的配对 表 12 ̄7 密码子与反密码子配对的摆动现象 tRNA 反密码子的第一位碱基 C A U G I 密码子的第三位碱基 G U A G C U U C A 3 参与蛋白质生物合成的酶和蛋白因子 (1) 主要的酶 1) 氨基酰 ̄tRNA 合成酶:位于胞液中ꎬ催化氨基 酸的活化ꎮ 2) 转肽酶:是核糖体大亚基的成分ꎬ催化 P 位与 A 位的氨基酸之间生成肽键ꎮ 3) 转位酶:其活性存在于延长因子 G 中ꎬ催化核 糖体的移位ꎮ (2) 蛋白质因子:见表 12 ̄ 8ꎮ 表 12 ̄ 8 参与蛋白质生物合成的各种蛋白因子 原核生物 真核生物 起始因子 3 种(IF ̄ 1、IF ̄ 2、 IF ̄ 3) 10 余 种 ( 至 少 9 种 elF1 ̄ 9) 延长因子 3种(EF ̄Tu、EF ̄Ts、 EF ̄G) 3 种 ( eEF1 ̄α、 eEF1 ̄ βγ、eEF ̄ 2) 释放因子 3 种(RF ̄ 1、RF ̄ 2、 RF ̄ 3) 1 种(eRF) (三) 蛋白质的生物合成过程 1 起始阶段 (1) 参与起始的因子:见表 12 ̄9ꎮ

第12章蛋白质的生物合成·129·% 表129蛋白质生物合成的起始因子 原核生物 直核生物 起始因子 生物学功能 起始因子 生物学功能 R-1 古据A位,防止其他RNA结合 elF-1 参与翻译的多个步骤 F.2 侵进Met-4RNA与小亚基结合 elF.2 促进Met-tRNA“与小亚基结合 IF-3 促进大,小亚基分离 clF.2B 结合小亚基,促进大,小亚基分离 F.3 结合小亚基,促进大,小亚基分离:使下4F mRNA与核糖体小亚基结合 elF-4A 有RNA螺旋酶活性,解开mRNA5'-端发夹结构 使其与小亚基结合 elF-4B 结合mRNA,促进其扫描定位起始AUC elF-4E 结合帽子结构 elF-4G 结合eF.4E、eF.3和PAB -5 促进各种因子脱落 elF-6 促进核糖体分离为大,小亚基 (2)起始的过程:见表12.10。 表1210蛋白质生物合成的起始过程 原核生物起始过程 真枝生物起始过程 具体过程 具体过程 ①核糖体大小亚F,与30s小亚基结合,E,占据A位,核糖①技糖体大小亚在F.6参与下,F.2B,F.3与小亚 基分离 体解离为50S大亚基和30S小亚基 基分离 基结合,核糖体解离为大亚基和小 亚基 ②mRNA在小亚①mRNA起始AUG上游s-D序列与小亚 ②Met-tRNAM cl.2与GP结合.然后Met-tRNA与 基上定位结合 其中16SRNA3'端的段序列百结 小亚基 F2结合,再结合到小亚基P位 合:②mRNA紧接SD序列后的短序列与 结 小亚基蛋白.1结合 3 Met-RNAN GTP.F,-RNA=复合物进人P位,对③mRNA在小亚①f.4E结合mRNA5”.子:②ohA 的结合 应在mRNA的起始密码AUG 基上位结合 结合蛋白结合mRNA3 结构和A尾再与核糖体小亚其 合:④F.4A消耗AP将mRNAS” 的一级结构解错使.RNA mRNA扫描,直到起始AUG与M tRNA“的反密码结合,mRNA最终在 小亚基正确定位 ④核糖体大亚基50s大亚基结合,同时GTP一GDP+5,释④核糖体大亚基60S大亚基结合,通过P.5的作用,同 结合 放F,F,和F:,形成70S起始复合物 结合 时GTP-GDP+P,释放各种因子 (3)S-D序列:也称核糖体结合位点(ibosomal点,小亚基16SRNA3'端有富含ccUcCU的短序列。 binding site.RBS).是指原核生物mRNA起始AUG密 与S.D序列配对」 码上游约8-13核苷酸部位,存在4-9个核苷酸组成 2.延长阶段 的一致性序列,富含AGGACG,是与小亚基的结合位 (1)参与延长的因子:见表1211
第 12 章 蛋白质的生物合成 129 表 12 ̄ 9 蛋白质生物合成的起始因子 原核生物 真核生物 起始因子 生物学功能 起始因子 生物学功能 IF ̄ 1 占据 A 位ꎬ防止其他 tRNA 结合 eIF ̄ 1 参与翻译的多个步骤 IF ̄ 2 促进 fMet ̄tRNA fMet与小亚基结合 eIF ̄ 2 促进 Met ̄tRNA Met i 与小亚基结合 IF ̄ 3 促进大、小亚基分离 eIF ̄ 2B 结合小亚基ꎬ促进大、小亚基分离 eIF ̄ 3 结合小亚基ꎬ 促进大、 小亚基分离ꎻ 使 eIF ̄ 4F  ̄ mRNA 与核糖体小亚基结合 eIF ̄ 4A 有RNA 螺旋酶活性ꎬ解开 mRNA5’ ̄端发夹结构ꎬ 使其与小亚基结合 eIF ̄ 4B 结合 mRNAꎬ促进其扫描定位起始 AUG eIF ̄ 4E 结合帽子结构 eIF ̄ 4G 结合 eIF ̄ 4E、 eIF ̄ 3 和 PAB eIF ̄ 5 促进各种因子脱落 eIF ̄ 6 促进核糖体分离为大、小亚基 (2) 起始的过程:见表 12 ̄ 10ꎮ 表 12 ̄ 10 蛋白质生物合成的起始过程 原核生物起始过程 真核生物起始过程 步骤 具体过程 步骤 具体过程 ①核糖体大小亚 基分离 IF3与 30S 小亚基结合ꎬIF1占据 A 位ꎬ核糖 体解离为 50S 大亚基和 30S 小亚基 ①核糖体大小亚 基分离 在eIF ̄ 6 参与下ꎬ eIF ̄ 2B、 eIF ̄ 3 与小亚 基结 合ꎬ 核 糖 体 解 离 为 大 亚 基 和 小 亚基 ②mRNA 在小亚 基上定位结合 ①mRNA 起始 AUG 上游 S ̄D 序列与小亚 基中 16S rRNA 3’端的一段序列互补结 合ꎻ②mRNA 紧接 SD 序列后的短序列与 小亚基蛋白 rps ̄ 1 结合 ② Met ̄tRNA Met i 与 小 亚 基 结合 eIF ̄ 2 与 GTP 结合ꎬ然后 Met ̄tRNA Met i 与 eIF ̄ 2 结合ꎬ再结合到小亚基 P 位 ③ fMet ̄tRNA fMet 的结合 GTP ̄IF2  ̄fMet ̄tRNA fMet复合物进入 P 位ꎬ对 应在 mRNA 的起始密码 AUG ③mRNA 在小亚 基上定位结合 ①eIF ̄ 4E 结合 mRNA5’ ̄帽子ꎻ②polyA 结合蛋白结合 mRNA3’ ̄polyA 尾巴ꎻ ③通过 eIF ̄ 4G 和 eIF ̄ 3 的介导ꎬ帽子 结构和 polyA 尾再与核糖体小亚基结 合ꎻ④eIF ̄ 4A 消耗 ATP 将 mRNA5’ ̄ 端的二级结构解链ꎬ使 Met ̄tRNA Met i 沿 mRNA 扫描ꎬ直到起始 AUG 与 Met ̄ tRNA Met i 的反密码结合ꎬmRNA 最终在 小亚基正确定位 ④核糖体大亚基 结合 50S 大亚基结合ꎬ同时 GTP→GDP +Piꎬ释 放 IF1 、IF2和 IF3 ꎬ形成 70S 起始复合物 ④核糖体大亚基 结合 60S 大亚基结合ꎬ通过 eIF ̄ 5 的作用ꎬ同 时 GTP→GDP+Piꎬ释放各种因子 ( 3) S ̄D 序列:也称核糖体结合位点( ribosomal binding siteꎬ RBS)ꎬ是指原核生物 mRNA 起始 AUG 密 码上游约 8~ 13 核苷酸部位ꎬ存在 4 ~ 9 个核苷酸组成 的一致性序列ꎬ富含 AGGAGGꎬ是与小亚基的结合位 点ꎬ小亚基 16S rRNA 3′ ̄端有富含 CCUCCU 的短序列ꎬ 与 S ̄D 序列配对ꎮ 2 延长阶段 (1) 参与延长的因子:见表 12 ̄ 11ꎮ

·130·生物化学与分子生物学学习纲要与同步练习 表12山蛋白质生物合成的延长因子 原核生物 真核生物 延长因子 生物学功能 延长因子 生物学功能 EF-Tu 与氨基酰-RNA及GTP结合,促进氨基 eEFl- 促使阳-RNA与枝糖体结合 酰-RNA进人A位 EF-Ts 结合EF.T,取代GD eEF1-By 使EFI-a再循环 EF-G 转位前活性,促进mRNA肤酰RNA由 EF.2转位 A位移至P位 (2)征长的过程.见表12.12 表1212蛋白质生物合成的延长 原核生物的延长 真核生物的延长 步骤 过程 ①进位 个氨基酰RNA按凰mRNA模板的指令进人并结合到核铝 体A位的过程 核糖体P位上起始氨基酰RNA的N甲酰甲硫 ②成 氢酰基或肽酰RNA的肱酰基转移到A位并与A位上氨基与原核生物类似,只是反应体系和延长 就-RNA的a一形成其庭的过程 因子不同,没有E位,卸载的RNA直 接从P位脱落 核使体向mRNA的3'端移动一个密码子的距离.使mRNA的 序列上的下一个密码子进人核蛋 3转位 白体的A位,而占据A位的肤酰RNA移人P位的过程,卸 我的BNA则移人E位,A位留空对应下一个密码 3.终止阶段 (1)参与终止的因子:见表12.13。 表1213蛋白质生物合成的终止因子 原枝生物 真生物 因子 生物学功能 因子 生物学功能 RF-1.RF-2.RF-3 识别终止信号,使大亚基转肽酶将“始 eRF 肽链释放 位”上已合成的多肽链水解释 (2)终止的过程:见表12.14。 表1214蛋白质生物合成的终止 原核生物终止过程 真核生物终止过程 ①识别终止密码,如BF,特异识别UAA,UAG:而RF,可识别UAA UGA:RF,刺激RF、F活性,协助释放。②当A位对应于 位,使P位上的肤酰 与原核生物相似,但只有一种释放因子,可识别所有 性变 从面使多肤 终止密码 、30S二个亚基,重新进入T 多肤链的合成。或变为单核糖材
130 生物化学与分子生物学学习纲要与同步练习 表 12 ̄ 11 蛋白质生物合成的延长因子 原核生物 真核生物 延长因子 生物学功能 延长因子 生物学功能 EF ̄Tu 与氨基酰 ̄tRNA 及 GTP 结合ꎬ促进氨基 酰 ̄tRNA 进入 A 位 eEF1 ̄α 促使 aa ̄tRNA 与核糖体结合 EF ̄Ts 结合 EF ̄Tuꎬ取代 GDP eEF1 ̄βγ 使 eEF1 ̄α 再循环 EF ̄G 转位酶活性ꎬ促进 mRNA ̄肽酰 ̄tRNA 由 A 位移至 P 位 eEF ̄ 2 转位 (2) 延长的过程:见表 12 ̄ 12ꎮ 表 12 ̄ 12 蛋白质生物合成的延长 原核生物的延长 真核生物的延长 步骤 过程 ①进位 一个氨基酰 ̄tRNA 按照 mRNA 模板的指令进入并结合到核糖 体 A 位的过程 核糖体 P 位上起始氨基酰 ̄tRNA 的 N ̄甲酰甲硫 ②成肽 氨酰基或肽酰 ̄tRNA 的肽酰基转移到 A 位并与 A 位上氨基 酰 ̄tRNA 的 α-氨基形成肽键的过程 核糖体向 mRNA 的 3′ ̄端移动一个密码子的距离ꎬ使 mRNA 的 序列上的下一个密码子进入核蛋 ③转位 白体的 A 位ꎬ而占据 A 位的肽酰 ̄tRNA 移入 P 位的过程ꎬ卸 载的 tRNA 则移入 E 位ꎬA 位留空对应下一个密码 与原核生物类似ꎬ只是反应体系和延长 因子不同ꎬ没有 E 位ꎬ卸载的 tRNA 直 接从 P 位脱落 3 终止阶段 (1) 参与终止的因子:见表 12 ̄ 13ꎮ 表 12 ̄ 13 蛋白质生物合成的终止因子 原核生物 真核生物 因子 生物学功能 因子 生物学功能 RF ̄ 1ꎬRF ̄ 2ꎬRF ̄ 3 识别终止信号ꎬ使大亚基转肽酶将“ 给 位”上已合成的多肽链水解释放 eRF 肽链释放 (2) 终止的过程:见表 12 ̄ 14ꎮ 表 12 ̄ 14 蛋白质生物合成的终止 原核生物终止过程 真核生物终止过程 ①识别终止密码ꎬ如 RF1 特异识别 UAA、UAGꎻ而 RF2 可识别 UAA、 UGAꎻRF3 刺 激 RF1 、 RF2 活 性ꎬ 协 助 释 放ꎮ ② 当 A 位 对 应 于 mRNA 的终止密码子时ꎬ终止因子作用于 A 位ꎬ使 P 位上的肽酰 基转移酶活性变为水解酶活性ꎬ从而使多肽链从核糖体上释放ꎬ同 时核糖体离开 mRNAꎬ解离为 50S、30S 二个亚基ꎬ重新进入下一条 多肽链的合成ꎮ 或变为单核糖体 与原核生物相似ꎬ但只有一种释放因子ꎬ可识别所有 终止密码