
第十一章 DNA的生物合成 学习目标 通过本章的学习,你应该能够: 掌握半保留复制的概念,原核生物(大肠杆菌)DNA聚合酶I的作用,解螺旋 酶,DNA拓扑异构酶、单链DNA结合蛋白、DNA连接酶、引物的作用: 逆转录概念和逆转录酶的作用;DNA突变的概念。 熟悉DNA复制所需的其它主要成分包括单链DNA模板、原料dNTP(dATP dGTP、dCTP、dTTP)等;原核生物DNA复制起始区的结构与功能:端粒的 概念、端粒酶的基本结构与作用:突变的主要类型,切除修复的基本过程。 了解遗传信息传递的中心法则,半保留复制的实验证据,DNA双向复制、复制 起始点、复制叉、复制子的概念;半不连续复制、前导链、后随链和冈崎片 段的概念;复制中引物、引发体的概念,DNA复制的高保真性及有关的机 制:大肠杆菌其它DNA聚合酶的作用,常见的真核生物DNA聚合酶及其 作用:原核生物复制的延长与终止时程,直核生物DNA复制时程与原核 生物的差异;端粒的概念及作用、端粒酶参与端粒合成的机制;滚环复制 等其他复制方式。反转录的生物学意义,常见的DNA损伤因素,突变的 后果与意义。DNA损伤修复的常见方式。 DNA是生物体或细胞内的主要遗传物质,其合成主要包括DNA复制、DNA修复合成和逆转录合成 DNA等过程。DNA复制(replication)是以亲代DNA分子为模板,按照碱基互补配对原则,在DNA聚合 酶等的作用下,沿5'一3'方向合成子代DNA分子的过程。DNA复制具有高度保真性,酶促修复系统可 以矫正复制中可能出现的错误。原核生物和真核生物DNA复制的基本原理和过程大致相似,但具体细 节上有所差别。对DNA复制机制的认识更多是来源于对原核生物的研究,真核生物的DNA复制过程 和参与的分子更为复杂和精致。 多种理化因素的作用及复制过程中出现的错误均可导致生物体D八A分子组成与结构发生变化,称 为DNA损伤(DNA damage),也称为突变。突变可分为碱基的错配、缺失、插人和重排等类型。在长期 的进化过程中,生物体细胞已形成了自己的DNA损伤修复系统,能及时纠正和修复损伤的DNA。DNA 修复的方式主要有直接修复、切除修复、重组修复和SOS修复等。本章主要讨论D八NA复制,逆转录以及 DNA的损伤及修复。 分

第十一章D以的生物合成 第一节DNA复制的基本特征 虽然不同生物基因组大小不同、结构也存在差异,复制上各有特点,但所有生物的基因组复制过程 都有一些基本特征。DNA复制的主要特征包括:半保留复制(semi-conservative replication)、双向复制 (bidirectional replication)和半不连续复制(semi-discontinuous replication)。DNA复制具有高保真性 (high fidelity)。 一、复制起始点核苷酸序列的特殊性 复制不是在基因组上的任何部位随机开始的,而是从DNA分子上的特定部位起始的,这一部位叫 做复制起始点(replication origin),常用on或o表示。DNA复制从起始点开始直到终点为止,每个这样 的DNA单位称为复制子或复制单元(replicon)。在原核细胞中,每个DNA分子只有一个复制起始点 因而只有一个复制子,而在真核生物中,DNA的复制是从许多起始点同时开始的,所以每个DNA分子 上有许多个复制子。 DNA复制起始点核苷酸序列有其结构上的特殊性,例如:大肠杆菌E.coi染色体DNA上有一个固 定的复制起始点,称为oiC,跨度为245bp,碱基序列分析发现这段DNA上游有3个13bp重复序列 (GATCTNTTNTTTT),下游有5个9bp重复序列[TT(A/T)TNCACC](图1I-1)。上游重复序列称为识 别区:下游重复序列碱基组成以A,T为主,称为富含AT(AT rich)区。这些部位是大肠杆菌中DnaA蛋 白识别、结合的位置。DNA双链中,AT间的配对只有2个氢键维系,故富含AT的区域容易发生解链 这些特殊的结构对于在DA复制起始过程中参与的酶和许多蛋白质分子的识别和结合都是必需的。 图111原核生物的复制起始部位 真核生物复制起始点比E.co的omC短。酵母DNA复制起始点含1Ibp富含AT的核心序列: A(T)TTTATA(G)TTTA(T),称为自主复制序列(autonomous replication sequence,ARS)。已发现的几个 多细胞真核生物DNA复制起点比细菌和酵母大,特异序列可能对复制起始并不重要,决定复制起始的 因素与核小体密度较低、靠近转录活性区有关。 二、复制的双向性 DNA双链从复制起始点向两个方向解开,复制沿两个方向同时进行,称为双向复制(bidirectional replication)。解开的两条模板单链和尚未解旋的DNA双链模板形成叉状结构,称为复制叉(replication fork),又称为生长叉(growing fork),如图11-2所示。含有一个复制起始点的一个完整DNA分子或DNA 分子上的某段区域被看做一个独立复制单元,称为复制子(r心plicon)。质粒、细菌染色体和噬菌体等通 常只有一个复制起始点,因而其DNA分子就构成一个复制子;真核生物染色体有多个复制起始点,所以 含有多个复制子。 三、复制的半保留性 在研究DNA复制方式的初期,人们提出了三种可能的方式,即全保留、半保留和混合式(图113) 最后,通过实验证实了DNA复制的方式是半保留复制
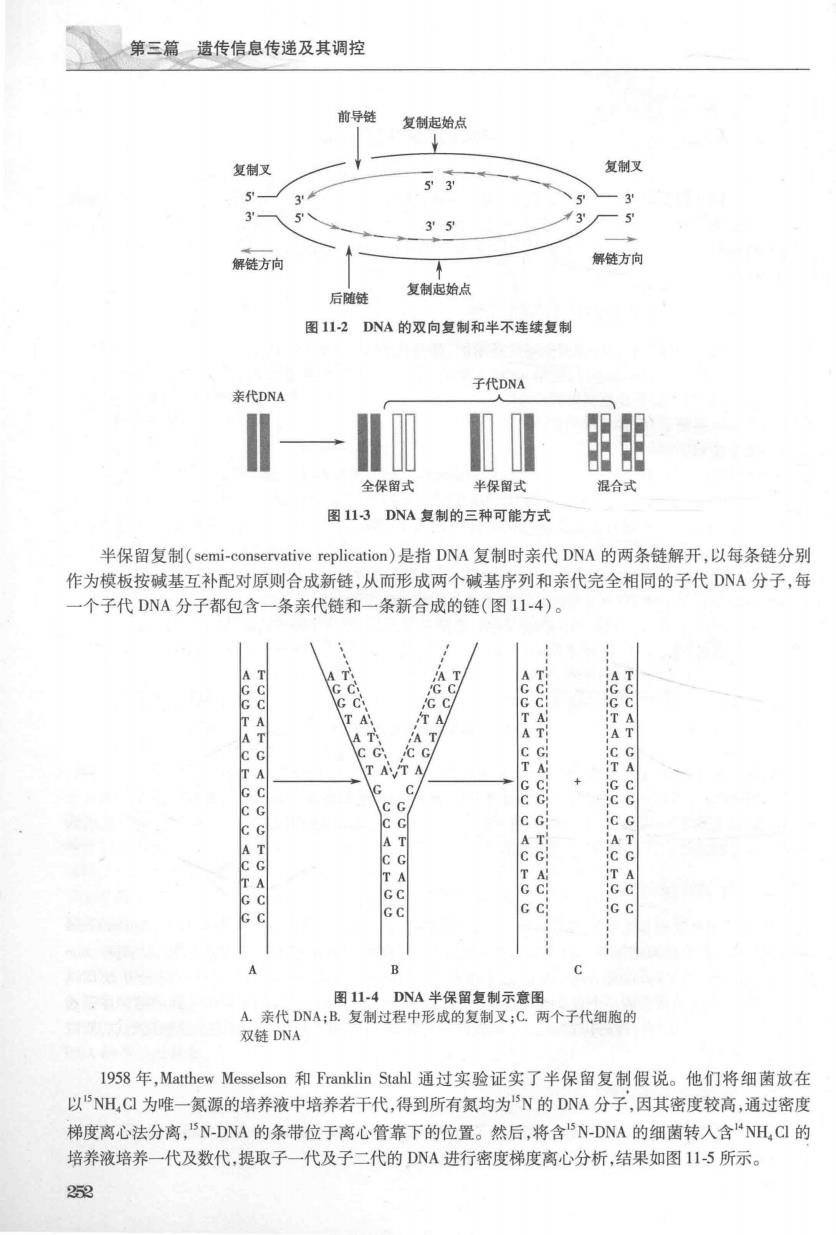
第三篇遗传信息传递及其调控 前导链 复鳞起始点 复制叉 复制又 5到 解链方向 解方向 后结 复制起始点 图1I2DNA的双向复制和半不连续复制 子代DNA 代DNA 图 全保留式 半保留式 混合式 图113DNA复制的三种可能方式 半保留复制(semi-conservative replication)是指DNA复制时亲代DNA的两条链解开,以每条链分别 作为模板按碱基互补配对原则合成新链,从而形成两个碱基序列和亲代完全相同的子代D八A分子,每 个子代DNA分子都包含一条亲代链和一条新合成的链(图114)。 c c e8 cc ic c A T 图114DNA半保留复制示意图 众代A复过客中形黄药定假C个于代的 l958年,Matthew Messelson和Franklin Stahl通过实验证实了半保留复制假说。他们将细菌放在 以“NH,C为唯一氯源的培养液中培养若干代,得到所有氮均为“N的DNA分子,因其密度较高,通过密度 梯度离心法分离,5N-DNA的条带位于离心管靠下的位置。然后,将含5N-DNA的细菌转人含“NH,C的 培养液培养一代及数代,提取子一代及子二代的DNA进行密度梯度离心分析,结果如图115所示。 25

第十一章DN的生物合成 川 含N-DNA的细睹 环装 第一代 日4 赞携养 第二代 器 图115证明DNA半保留复制的经典实验 实验结果显示,培养一代后的DNA分子其密度介于5N-DNA和N-DNA之间,说明复制产生的两 个DNA分子中都有一条链是N-DNA单链,另一条是“N-DNA单链,即杂合的DNA。培养第二代,得到 等量的杂合DNA和N-DNA。继续培养,杂合DNA的含量呈几何级数减少。这一实验结果符合半保留 复制的假说,证实了DNA的复制方式为半保留复制。这种复制方式的意义在于,能使亲代DNA所含的 信息以极高的准确度传递给子代DNA分子,体现了生物遗传过程的相对保守性。 四、复制的半不连续性 DNA分子在复制过程中会产生两条DNA新链,其合成方向是5'3'。新链和模板链之间是反向平 行的关系,所以在复制过程中一条新链的合成方向与复制叉前进的方向相同,能连续合成:而另一条新 链的合成方向与复制叉前进方向相反,合成是不连续、分段进行的,这种复制方式称为半不连续复制 (semi-)。能连续合成的链称为前导链(leading strand),不能连续合成的链称为 后随链(lagging strand),见图1l-2。后随链在合成过程中必须待模板DNA解开足够的长度才能合成新 链,所以其复制方式是先合成一些短的DNA片段,然后再通过连接酶将其连接形成完整的长链。1968 年,日本学者Reiji Okazaki利用电子显微镜和放射自显影技术观察到了后随链复制过程中有多个小片 段的生成,后人将其命名为冈崎片段(Okazaki fragment)。真核生物中冈崎片段的长度约为100~ 200bp,而原核生物约为1000-2000bp。 五、复制需要RNA引物 DNA复制起始过程需要先合成引物(primer),引物是由引物酶催化合成的短链RNA分子。母链 DNA解成单链后,不会立即按照模板序列将dNTP聚合为子链DNA.这是因为依赖DNA的DNA聚合 酶(DNA-dependent DNA polymerase),简称DNA聚合酶(DNA pol),只能催化核酸片段的3'-OH末端与 NTP之间的聚合,不具备催化两个游离dNTP之间形成磷酸二酯键的能力。因此,复制起始部位合成 的引物只能是RNA,引物酶属于RNA聚合酶。短链引物RNA为DNA的合成提供3'-OH末端,在DNA 聚合酶催化下逐一加入dNTP而形成DNA子链。 六、复制的高度保真性 DNA复制的保真性是遗传信息稳定传代的保证。生物体内至少有3种机制实现保真性:①遵守严 格的碱基配对规律;②DNA聚合酶对碱基的选择功能;③复制出错时D八A聚合酶有即时的校对功能。 (一】复制的保直性依输正确的碱基洗怪 DNA复制保真的关键是正确的碱基配对,而碱基配对的关键又在于氢键的形成。G和C以三个氢 键,A和T以2个氢键维持配对,错配碱基之间难以形成氢键。除化学结构限制外,DNA聚合酶对碱基 253

第三篇遗传信息传递及其调控 具有选择作用。 DNA聚合酶Ⅲ(DNA polⅢ)是在原核生物DNA链延长中起催化作用的酶。利用“错配”实验发 现,DNA polⅢ对核苷酸的掺入具有选择功能。例如,用21聚脱氧腺苷酸(poly(dA)21)作模板,用poly (T)20作复制引物,可以观察引物3'-0H端连上的是否为胸苷酸(T)。尽管反应体系中4种核苷酸都 存在,第21位也只会出现T。但若只向反应体系加单一的NTP作底物,就“迫使”引物在第21位延长 中出现错配。用柱层析技术可以把DNA polⅢ各个亚基组分分离,然后又再重新组合。如果重新组合 的D八AplⅢ不含亚基,复制错配率出现较高,说明e亚基是执行碱基选择功能的。 嘌吟的化学结构能形成顺热和反式构型,与相应的感啶形成氢罐配对,原吟应处干反式构型。而要 形成嘌呤-嘌呤配对,则其中一个嘌吟必须旋转180°,形成反式构型。DNA polⅢ对嘌吟的不同构型表 现不同的亲和力,因而实现其选择功能。 DNApolⅢ有10个亚基,以a、e和0作为核心酶并 组成较大的不对称二聚体。核心酶中,α亚基有5'→3' ONA Pol I 聚合酯活性,有3'5核酸外切酶活性及减基洗择功 外切活性案合活性 能。未发现日亚基有催化活性,可能起维系二聚体的作 用。对各亚基功能的深人研究认为:在核苷酸聚合之前 dCTP 或聚合当时,酶就可以控制碱基的正确选择。 【二)聚合酶中的核酸外切酶活性在复制中辨认切 除错配碱基并加以校正 无活性 原核生物的DNA pol I、真核生物的DNA pol8和 5 DNA polε的3'-5核酸外切酶活性都很强,可以在复制 过程中辨认并切除错配的碱基,对复制错误进行校正,此 B 过程又称错配修复(mismatch repair) 以DNA pol I为例(图11-6),图中的模板链是G,新 确配对 链错配成A而不是C。DNA pol I的3'→5核酸外切酶 活性就把错配的A水解下来,同时利用5'一3聚合酶活性补回正确配对的C,复制可以继续下去,这种 功能称为校对(proofreading)。实验也证明:如果是正确的配对,3'→5'核酸外切酶活性是不表现的。 DNAp1I还有5-3'外切酶活性,实施切除引物、切除突变片段的功能。 第二节参与DNA复制的酶和蛋白质因子 目前,在大肠杆菌中发现的与DNA复制相关的蛋白质大约有30种,在真核生物中相关的蛋白质更 多。主要有DNA聚合酶、拓扑异构酶、解螺旋酶、单链DNA结合蛋白、引物酶和DNA连接酶。 一、DNA聚合酶 DNA聚合酶全称是依赖DNA的DNA聚合酶(DNA-dependent DNA polymerase,DDDP),简称DNA pol,是1958年由Arthur Kornberg在大肠杆菌中首次发现。目前在原核及真核生物中都发现了多种类型 的DNA聚合酶,它们主要表现出以下三种催化活性:①5'→3'方向的聚合酶活性,催化3',5'-磷酸二酯 键的形成,使DA链沿5一3'方向延长;②5'一→3'核酸外切酶活性,能从5'一→3'方向水解核酸单链,在 DNA复制中主要用于对引物的水解;③3'一→5'核酸外切酶活性,能从3'→5'方向将复制过程中错配的核 苷酸水解,具有校正修复的功能 原核生物的DNA聚合酶分为三型(表II-1):DNA pol I、DNA pol II和DNA polⅢ。DNA pol I具 有上述三种催化活性,在DNA复制过程中主要用于填补引物切除后留下的空隙。DNA pol【由一条含 254