
第十六章细胞内膜系统 学习目标 通过本章学习,你应该能够: 掌握内膜系统概念、内质网、高尔基体、溶酶体、过氧化物酶体结构功能等 熟悉内质网、高尔基体、溶酶体的结构、功能等。 了解内膜系统各细胞器异常与疾病的关系。 在直核细胞的细胞质中,有一些由膜构成的细胞器,它们被此相互关联,组成了一个精密而复杂的 膜系统,把细胞质分隔成了多个特异性的功能区,而每个功能区均含有自身特异性的酶和其他特定的成 分。通常我们把细胞质中由核膜、内质网膜和高尔基复合体膜的小管、小泡和扁囊组成的系统称为内膜 系统(endomembrane system)。内膜系统和质膜是相连的,同时和质膜一样,也是一种动态结构,具有流 动性、镶嵌性、不对称性和蛋白质极性。各种内膜之间可通过出芽和融合的方式进行交流,因此,溶酶 体、过氧化物酶体和分泌泡均可看做是内膜系统的衍生物。内膜系统不含有DNA,其功能活动和装配 完全受核DNA的控制。线粒体由于含有自身DNA,其功能活动和装配要受到核DNA和自身DNA的共 同调控,属半自主性细胞器。 内膜系统是真核细胞完成各种复杂生命活动所必需的基本结构,也是不同于原核细胞的重要特征。 真核细胞中,内膜系统把细胞质分隔为内膜系统的腔区和胞质溶胶区,而内膜系统中的各种成分又分为 更加精细的区域,从而使细胞质出现了高度区域化(compartmentalization)。 本章将着重描述内质网、高尔基复合体、溶酶体以及过氧化物酶体的结构、功能和病变。核膜部分 将在相关章节叙述。 第一节内质网 内质网(endoplasmic reticulum,ER)的概念是随着电子显微镜技术的发展而建立的。945年 KRP:用电镜观察培养的小鼠成纤维细胞时,发现位于细胞质内靠近细胞核的内质区分布着一些 小管、小毒连成的网结构,故称为内质网(endoplasmic reticulum,ER,这-名称一直沿用至今。 内质网广泛分布于除成熟红细胞以外的所有真核细胞的细胞质中,虽然体积仅占细胞总体积的 10%,但其面积却占整个细胞生物膜系统的50%以上。内质网是由单位膜围绕形成的膜性结构,形态 呈泡状、管状和扁囊状,它们相互连通形成封闭管道系统,并常与外层核膜相连续,内质网腔与核周间隙 相通。研究表明,内质网在蛋白质和脂类合成等方面具有非常重要的作用。 一、内质网的类型与形态结构 内质网是细胞内一个精细的膜囊结构,由内质网膜相互连接形成一个连续的内腔相通的膜性管道 888
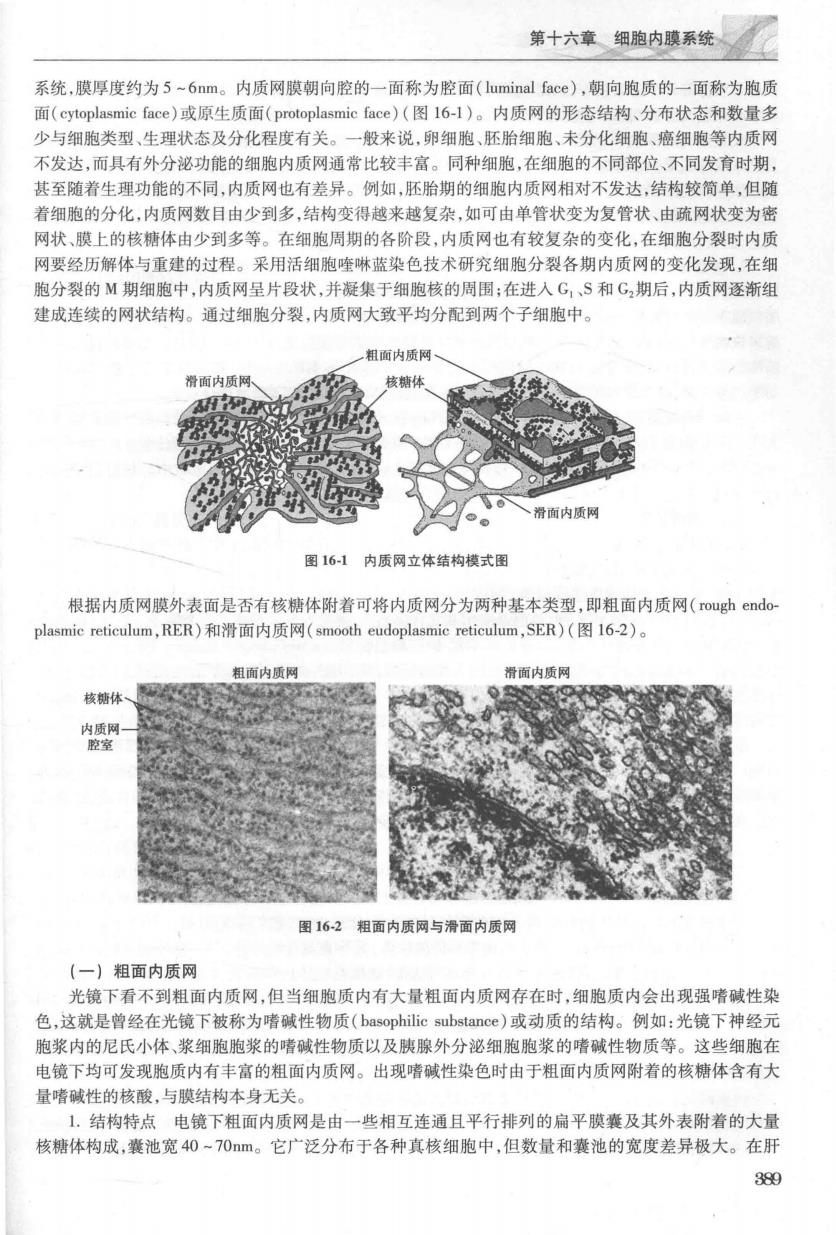
第十六幽内速系统 系统,膜厚度约为5-6nm。内质网膜朝向腔的一面称为腔面(luminal face),朝向胞质的一面称为胞质 面(cytoplasmic face)或原生质面(protoplasmic face)(图16-l)。内质网的形态结构、分布状态和数量多 少与细胞类型、生理状态及分化程度有关。一般来说,卵细胞、胚胎细胞、未分化细胞、癌细胞等内质网 不发达,而具有外分泌功能的细胞内质网通常比较丰富。同种细胞,在细胞的不同部位、不同发育时期 其至随着生理功能的不同,内质网也有差异。例如,胚胎期的细胞内质网相对不发达,结构较简单,但随 著细胞的分化,内质网数目由少到多,结构弯得战来城复杂,如可由单管状变为复管状、由疏网状变为密 网状、膜上的核糖体由少到多等。在细胞周期的各阶段,内质网也有较复杂的变化,在细胞分裂时内质 网要经历解体与重建的过程。采用活细胞喹啉蓝染色技术研究细胞分裂各期内质网的变化发现,在细 胞分裂的M期细胞中,内质网呈片段状,并凝集于细胞核的周围;在进人G,、S和G,期后,内质网逐渐组 建成连续的网状结构。通过细跑分裂,内质网大致平均分配到两个子细胞中。 一粗面内质 滑面内质网 核糖体 滑面内质网 图16-1内质网立体结构模式图 根据内质网膜外表面是否有核糖体附着可将内质网分为两种基本类型,即粗面内质网(rough endo plasmic reticulum,RER)和滑面内质网(smooth eudoplasmie reticulum,SER)(图16-2)。 粗面内质网 滑面内质网 核糖体 图162粗面内质网与滑面内质网 (一)粗面内质网 光镜下看不到粗面内质网,但当细胞质内有大量粗面内质网存在时,细胞质内会出现强嗜碱性染 色,文就是曾经在光镜下被称为嗜碱性物质(basophilic substance)或动质的结构。例如:光镜下神经元 胞浆内的尼氏小体、浆细胞胞浆的嗜碱性物质以及胰腺外分泌细胞胞浆的嗜碱性物质等。这些细胞名 电镜下均可发现胞质内有丰富的粗面内质网。出现嗜碱性染色时由于粗面内质网附着的核糖体含有大 量者碱性的核酸与草结构本身无关 1.结构特点电镜下粗面内质网是由一些相互连通且平行排列的扁平膜囊及其外表附着的大量 核糖体构成,囊池宽40~70m。它广泛分布于各种真核细胞中,但数量和囊池的宽度差异极大。在肝 39g

第四篇细胞的结构与功能 细胞和胰腺等外分泌细胞中,粗面内质网囊池很狭窄,囊池内中空或含有少量中等电子密度物质:而在 成纤维细胞及滑膜细胞中,粗面内质网囊池比较宽大 大量研究结果表明,相面内质网发秋程度可作为判新细胞分化程度和功能状态的一项重要指标 凡是未成熟或未分化细胞与相应的正常成熟细胞相比,其粗面内质网不发达,如干细胞、各种母细胞利 胚胎细胞、培养细胞。这些细胞生长快速,细胞质中有丰富的游离核糖体,主要合成内源性蛋白质以供 自身生长、分裂需要。而执行分泌功能的细胞,如胰腺腺泡上皮细胞、肝细胞和浆细胞等,其内质网比较 发达,但在分泌活动旺感或低下时,内质网也有数量多或少的变化 2.核糖体核糖体(0soe)是一种非膜性结构的生物大分子复合物.是细胞内蛋白质合成的场 所,普遍存在于原核细胞和真核细胞内。真核细胞内核糖体包括两种类型:一种是附着在内质网膜和 层核膜表面的核糖体,称附着核糖体(attached ribosome):另一种是游离在细胞质基质中的核糖体,称为 游离核糖体(f©e ribosome)。附着核糖体和游离核糖体在执行蛋白质合成功能过程中,都不是以单个核 糖体的形式进行的,而是由RNA链把多个甚至几十个核糖体串联在一一起,形成具有合成蛋白质特殊 功能的聚合体,即多聚核糖体(polyribosome),多聚核糖体极大地提高了蛋白质合成的速率。 电镜下观察显示,核糖体是一种直径15 25m的小颗粒,由大小两个亚基构成。真核细胞核糖体 大亚基为60S,含有5 S rRNA,5.8 S rRNA,28SRNA等3种类型rRNA和49种蛋白质:小亚基为40S,由 一条18SRNA和33种蛋白质构成。核糖体通常以大、小亚基的形式存在于细胞质中。只有当细胞在 合成蛋白质时,大、小亚基才结合在一起,蛋白质合成结束,大、小亚基又解离。 (二】滑面内质网 滑面内质网也同样广泛存在于各种细胞内的细胞器,但在形态与功能上都不同于粗面内质网。滑 面内质网由分支的管道形成较为复杂的立体结构,膜表面没有核糖体附着,因表面光滑而得名。多为分 支的小管,很少扩大形成小泡或扁囊。小管直径约为50~100nm,膜较薄约4-5m,它们相互连接成 网,在一定部位可与粗面内质网、核膜及高尔基复合体相车,偶见与质膜相连。在光镜下,由于细胞基质 和沿面内质网皆为老酸性物质,所以在苏木精伊红边伍法(hematoxylin-eosin staining,HE)热色切片下 不宜识别。 一般来说,细胞质内滑面内质网丰富的细胞,其嗜酸性较强,如:胃壁细胞、寥丸间质细胞、黄 体细胞和肾上腺皮质细胞等。有时,滑面内质网和集中于细胞质的某一局部,光镜下呈粉红色小体,常 称为嗜酸小体(body)。在病毒性肝炎的肝细胞中即可见到 滑面内质网的膜比粗面内质网的膜薄,电镜下不易分辨出单位膜的三层结构。构成滑面内质网膜 的蛋白质及脂类处于不断的代谢更新之中。同位素实验表明,H标记的氨基酸和脂类首先渗入粗面内 质网膜中,然后出现在滑面内质网的膜上。表明新生的膜蛋白和膜磷脂主要由粗面内质网合成,经连接 部位移向滑面内质网。而滑面内质网本身也可合成部分磷脂 二、内质网膜的化学组成 粗面内质网和滑面内质网可用物理方法分离。将组织或细胞匀浆后,其内质网可破碎成许多碎片 采用密度梯度离心技术可将这些内质网的碎片从细胞中分离出来,它们多是直径为100m的封闭小 囊,称为微粒体(nicrosome)。来自粗面内质网的微粒体,其外表面有核糖体附着,称粗面微粒体(rouh microsomes)。由于核糖体总是存在于微粒体的外表面,说明微粒体内部在生化上相当于内质网腔。在 匀浆中还有些微粒体大小与粗面微粒体相仿,但表面没有核糖体附着,称滑面微粒体(smooth micro soms)。滑面微粒体一部分来自滑面内质网,一部分可能来自细胞膜、高尔基体或其他细胞器的碎片 因此滑面微粒体的成分比较复杂。但在肝细胞中光面内质网丰富,因此肝匀浆中多数滑面微粒体来自 滑面内质网。 内质网化学组成方面的资料主要来源于对微粒体膜的研究。内质网膜是由脂类和蛋白质组成。内 质网膜含30%~40%脂类和60%~70%蛋白质,脂类成分中主要为磷脂,此外还有中性脂肪、磷脂酰肌 醇、缩醛磷脂和一些神经节苷脂等。各类磷脂的比例大致为:磷脂酰胆碱占55%,磷脂酰乙醇胺为 390

第十六章细胞内膜系统 209%~25%,磷脂酰丝氨酸5%~10%,磷脂酸肌醇5%~10%,鞘磷脂4%~7% 内质网的蛋白质含量比细胞膜多。这些蛋白质既有边周蛋白,也有整合膜蛋白,它们在膜上呈 不对称分布,有的偏向腔面,有的偏向胞质面。蛋白质成分中多为糖蛋白,其中一些为酶。内质网膜 中约含有30~40种酶,其中葡萄糖-6-磷酸酶(glucose-6-phosphatase,G-6-P)被视为内质网膜的标志 酶,另一类标志酶是电子传递体系。如:NADH细胞色素c还原酶、NADH细胞色素b5还原酶、细胞 色素b5、细胞色素P450等。细胞色素P450主要位于滑面内质网的胞质面,是滑面内质网特有的标 记酶。 三、内质网的功能 内质网是细胞内多功能细胞器,除了进行蛋白质合成、脂类合成、糖代谢和解毒作用之外,还与 物质运输、蛋白质折叠修饰以及为细胞提供物理支撑等密切相关。粗面内质网和滑面内质网在结构 上既有共性又存在差异。除了是否存在核糖体附着以外,两者在蛋白质种类和数量上,特别是酶系 统方面存在较大的差别。因此,两者在功能上存在差异。目前已知至少有两种膜蛋白是粗面内质网 所特有的,并与核糖体与粗面内质网的结合有关,这两种蛋白质称为核糖体结合糖蛋白(ibophorin) I和Ⅱ,其分子量分别为63kD和65kD,它们是膜的整合糖蛋白。另外,粗面内质网膜中还含有信号 识别颗粒的受体(signal-recognition partice receptor)等蛋白质,而滑面内质网中则没有上述这些蛋白 质的存在。 (一)粗面内质网的功能 1.粗面内质网与蛋白质合成由于粗面内质网与核糖体结合,且含有丰富的酶类,使得其功能呈 现多样化。粗面内质网主要参与合成分泌性蛋白质、膜整合蛋白质、内质网驻留蛋白(endoplasmic retic )及溶酶体蛋白等,其中分泌蛋白包括各种肽类激素、酶类和抗体等。由内质网合成 的分泌蛋白质首先会释放进入内质网腔室进行初步加工,然后输送到高尔基复合体进行进一步的加工、 修饰,最终包装成含有成熟分泌颗粒的分泌泡,再由细胞表面释放出去。由粗面内质网参与合成的分泌 蛋白,膜整合蛋白、驻留蛋白及溶酶体蛋白等,多数都需要在内质网腔室进行初步的糖基化修饰后,被转 运至相应的部位 核糖体是蛋白质合成的场所,胞质溶胶中的游离核糖体与mRNA结合以后形成的多聚核糖体是蛋 白质合成的开始。游离多聚核糖体与附着于内质网膜的多聚核糖体在结构上没有任何区别,但是游离 的多聚核糖体合成的蛋白质不会进入粗面内质网,而附着的多聚核糖体合成的肽链将进入粗面内质网 腔,其关键在于结合的mRNA不同。粗面内质网上蛋白质合成的过程遵循信号假说(signal hypothesis), 信号假说由G.Blobel和D.Sabatini等在1975年提出,该假说认为蛋白质N端序列可以作为信号肽,负 责指导蛋白质转运至内质网上完成合成,详细阐述了粗面内质网上合成蛋白质的生物学过程。 G.Blobel和D.Sabatini也因为提出这一假说而获得1999年诺贝尔生理学或医学奖。 (I)信号假说(signal hypothesis):蛋白质合成是在mRNA的指导下完成的。研究表明,所有在相 面内质网合成的蛋白质,其连接多聚核糖体的mRNA在起始端均有一段编码约18~30个氨基酸的信号 序列(signal sequence)。它们位于起始密码子(AUG)之后。由信号序列翻译出的肽链称为信号肽 (signal peptide),特指蛋白质多肽链上的一段连续的特定氨基酸序列,具有分选信号的功能。可位于多 肽链的任何部位,完成分选任务后常被切除。除了信号肽以外,还有一种信号斑(signal patch),指位于 多肽链不同部位的几个特定氨基酸序列经折叠后形成的斑块区,具有分选信号的功能。信号斑是一种 三维结构。完成分选任务后仍然存在(图16-3)。 当核糖体与mRNA起始端结合以后首先翻译出这段信号肽,这时,胞质内的信号识别颗粒(signal recognition particle,SRP)可以识别新生信号肽并与之结合形成SRP-核糖体复合体。SRP是由6个多时 亚单位和一个小的7SRNA(图16-4)组成的RNA-蛋白质复合体。SRP有两个功能域,一个功能域用以 识别信号肽,结合含有疏水核心的信号肽使其不能折叠而能穿越内质网:另一个功能域干扰氨基酰 391
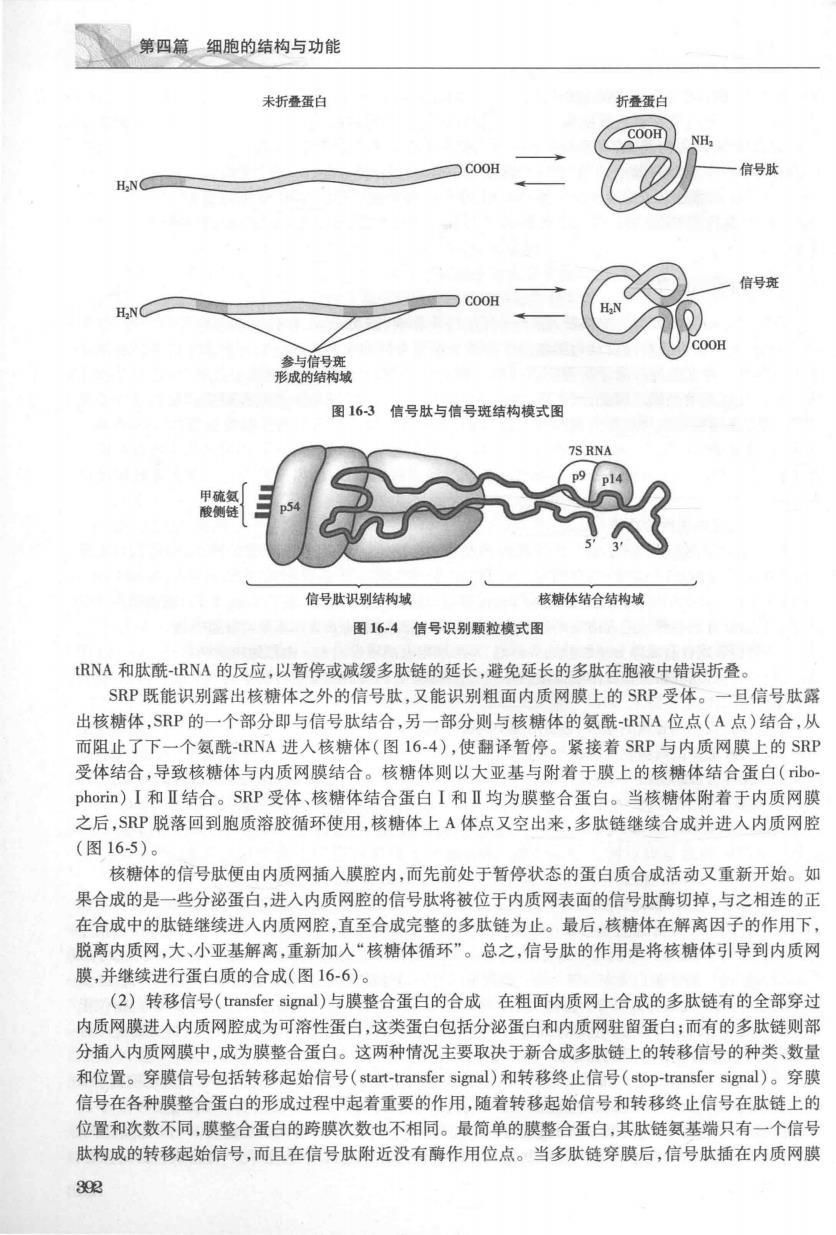
第四篇细胞的结构与功能 未折叠蛋白 折叠蛋白 信号 H.N 图16-3信号肽与信号斑结构模式图 信号肽识别结构域 核糖体结合结构城 图16-4信号识别颗粒模式图 RNA和肽酰-RNA的反应,以暂停或减缓多肽链的延长,避免延长的多肽在胞液中错误折叠。 SRP既能识别露出核糖体之外的信号肽,又能识别粗面内质网膜上的SRP受体」 一且信号肽露 出核糖体,SRP的一个部分即与信号肽结合,另一部分则与核糖体的氨酰-RNA位点(A点)结合,从 而阻止了下一个氨酰-RNA进入核糖体(图16-4),使翻译暂停。紧接着SRP与内质网膜上的SRP 受体结合,导致核糖体与内质网膜结合。核糖体则以大亚基与附着于膜上的核糖体结合蛋白(b0 pon)I和Ⅱ结合。SRP受体、核糖体结合蛋白I和Ⅱ均为膜整合蛋白。当核糖体附着于内质网膜 之后,SP脱落回到胞质溶胶循环使用,核糖体上A体点又空出来,多肽链继续合成并进入内质网脑 (图16-5)。 核糖体的信号肽便由内质网插入膜腔内,而先前处于暂停状态的蛋白质合成活动又重新开始。如 果合成的是一些分泌蛋白,进入内质网腔的信号肽将被位于内质网表面的信号肽酶切掉,与之相连的正 在合成中的肽链继续进入内质网腔,直至合成完整的多肽链为止。最后,核糖体在解离因子的作用下 脱离内质网,大、小亚基解离,重新加入“核糖体循环”。总之,信号肽的作用是将核糖体引导到内质网 膜,并继续进行蛋白质的合成(图16-6)。 (2)转移信号(ransfer signal)与膜整合蛋白的合成在粗面内质网上合成的多肽链有的全部穿起 内质网膜进入内质网腔成为可溶性蛋白,这类蛋白包括分泌蛋白和内质网驻留蛋白;而有的多肽链则部 分插人内质网膜中,成为膜整合蛋白。这两种情况主要取决于新合成多肽链上的转移信号的种类、数量 和位置。穿膜信号包括转移起始信号(start-transfer signal)和转移终止信号(stop-transfer signal)。穿膜 信号在各种膜整合蛋白的形成过程中起着重要的作用,随着转移起始信号和转移终止信号在肽链上的 位置和次数不同,膜整合蛋白的跨膜次数也不相同。最简单的膜整合蛋白,其肽链氨基端只有一个信号 肽构成的转移起始信号,而且在信号肽附近没有酶作用位点。当多肽链穿膜后,信号肽插在内质网膜 302