
第三篇遗传信息传递及其调控 核心 三启动子区 二闭合的启动子复合数 二开放的启动子复合物 三流产性起好 延长 。因子释 图124原核生物基因的转录起始 (三)真核生物基因启动子的特征 与原核生物相比,真核生物的启动子区域更为复杂。如前所述,真核生物有三种RNA聚合酶,分别 转录不同类型的基因。因此,可据此将真核生物基因的启动子分为三类,即I型、Ⅱ型和Ⅲ型启动子,分 别对应于RNA聚合酶I、Ⅱ和Ⅲ。这三种类型启动子的特征各不相同,其中以RNA聚合酶Ⅱ的启动子 即Ⅱ型启动子最为复杂(本章第三节,第14章)。 第二节转录的基本过程 转录过程可大致分为三个阶段:起始、延长和终止。原核生物与真核生物转录相比,其转录过程既 有相似的地方,也有很多不同之处。下面分别予以介绍。 一、原核生物转录的基本过程 (一)起始阶段 转录起始阶段的关键是RNA聚合酶识别并结合待转录基因的启动子从而启动转录,这也正是转录 调控的关键步骤。起始阶段可分为以下三个步骤。 第一步,闭合复合体(closed complex)形成。g因子与核心酶结合组装为RNA聚合酶全酶,在DNA 278
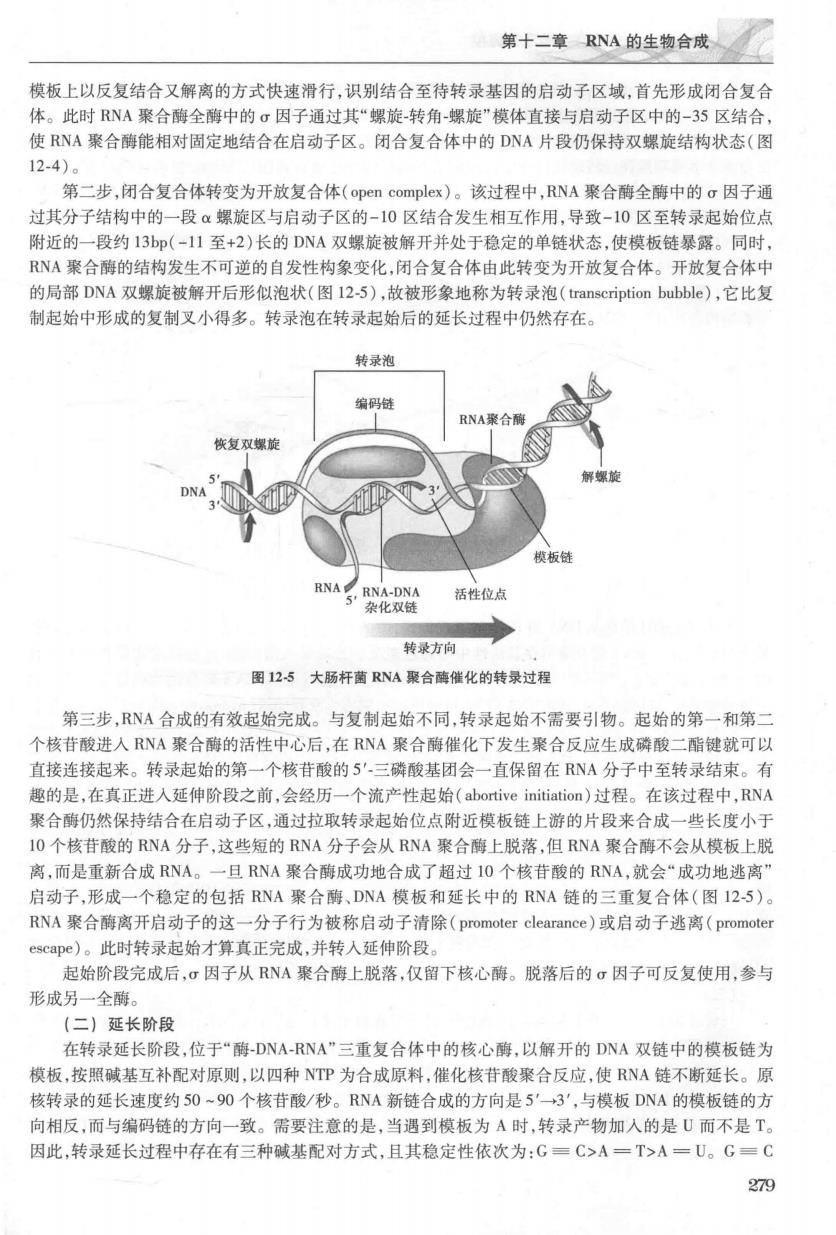
第十二重R认的生物合成☑ 模板上以反复结合又解离的方式快速滑行,识别结合至待转录基因的启动子区域,首先形成闭合复合 体。此时RNA聚合酶全酶中的行因子桶时其“摆旋转角辄旋”模体直接与启动子区中的一35区结合 使RNA聚合酶能相对固定地结合在启动子区。闭合复合体中的DNA片段仍保持双螺旋结构状态(图 12-4)。 第二步,闭合复合体转变为开放复合体(open complex)。该过程中,RNA聚合酶全酶中的c因子通 过其分子结构中的一段α螺旋区与启动子区的-10区结合发生相互作用,导致-10区至转录起始位点 附近的一段约13n(-11至+2)长的DNA双提旅被解开并处干稳定的单鞋状态,使模板韩暴碳。同时 RNA聚合酶的结构发生不可逆的自发性构象变化,闭合复合体由此转变为开放复合体。开放复合体中 的局部DNA双螺旋被解开后形似泡状(图12-5),故被形象地称为转录泡(transcription bubble),它比复 制起始中形成的复制叉小得多。转录泡在转录起始后的延长过程中仍然存在。 转录泡 NA蜜合 复 模板创 活性位点 转录方向 图125大肠杆菌RNA聚合酶催化的转录过程 第三步,RNA合成的有效起始完成。与复制起始不同,转录起始不需要引物。起始的第一和第二 个核苷酸进入RNA聚合酶的活性中心后,在RNA聚合酶催化下发生聚合反应生成磷酸二酯键就可以 直接连接起来。转录起始的第一个核苷酸的5'三磷酸基团会一直保留在RNA分子中至转录结束。有 趣的是,在真正进人延伸阶段之前,会经历一个流产性起始(abortive initiation)过程。在该过程中,RNA 聚合酶仍然保持结合在启动子区,通过拉取转录起始位点附近模板链上游的片段来合成一些长度小于 IO个核苷酸的RNA分子,这些短的RNA分子会从RNA聚合酶上脱落,但RNA聚合酶不会从模板上脱 离,而是重新合成RNA。一日RNA聚合酶成功地合成了招时10个核苷酸的RNA,就会“成功地逃离 启动子,形成一个稳定的包括RNA聚合酶、DNA模板和延长中的RNA链的三重复合体(图125)。 RNA聚合酶离开启动子的这一分子行为被称启动子清除(promoter clearance)或启动子逃离(promoter escap心)。此时转录起始才算真正完成,并转入延伸阶段 起始阶段完成后,σ因子从RNA聚合酶上脱落,仅留下核心酶。脱落后的▣因子可反复使用,参与 形成另一全酶。 (二)延长阶段 在转录延长阶段,位于“酶-DNA-RNA”三重复合体中的核心酶,以解开的DNA双链中的模板链为 模板,按照碱基互补配对原则,以四种NTP为合成原料,催化核苷酸聚合反应,使RNA链不断延长。原 核转录的延长速度约50~90个核苷酸/秒。RNA新链合成的方向是5'一3',与模板DNA的模板链的方 279

第三篇遗传信息传递及其调控 配对有三个氢键,最稳定:A=T配对主要在DNA双链形成,A一U配对只在DNA-RNA杂化双链或 RNA分子上形成,是三种配对中稳定性最低的 在转录过程中,转录泡前方的DNA不斯进入转录复合体并被解链,而转录复合体通过后,DNA双 链会重新形成双螺旋。转录复合体覆盖约3Obp的DNA,DNA解链范围约I7bp。而新合成的RNA链又 和DNA双链中的模板链配对形成长约8bp的RNA-DNA杂化双链。随着RNA链的不断合成延长,RN 的5'端部分则不断脱离模板链并向转录复合体外伸展。 在电子显微镜下观察原核生物的转录可以看到,在同一DNA模板上,有多个转录过程同时在进行 并且一条mRNA链上也有多个核糖体正在进行下一步的蛋白质合成过程,可见转录尚未完成,翻译已 在进行(图12-6)。因此,对于原核生物来讲,转录和翻译是紧密偶联,几乎同时进行的。而真核细胞有 核膜结构存在,故而将转录和翻译分隔在细胞核和细胞质中进行。 DNA 核糖体 图126原核生物转录与翻译同时进行 为了保证贵传信息从DNA到RNA的准确传递,RNA聚合在转录时也会对证长的新鞋行使两利 校读功能。第一,RNA聚合酶可在其活性中心通过逆反应重新引入焦磷酸,从而移除错误摻入的核苷 酸,这种校读机制称为焦磷酸解编辑(pyrophosphorolytic editing);第二,RNA聚合酶可以后退一个或多 个核苷酸单位,切除包含错误的RNA序列,这种校读机制称为水解编辑(hydrolytic editing)。然而,尽管 有上述校读机制,转录的精确度(参入1万个核苷酸出现1个错误)仍远低于复制的精确度(摻入1千万 个核苷酸出现1个错误)。因为转录合成的RNA仅仅旨在“抄录”遗传物质DNA中的序列信息,一个基 因可以转录产生许多RNA拷贝,而且RNA在细胞内很快会被降解和替代,所以转录产生错误RNA对 细胞的影响远比复制产生错误DNA对细胞的影响小。 放线菌素D(actinomycin d)是一种源于十寒的裤露菊属细莉中分离出来的放线黄素类多肽类抗出 素,能抑制原核生物和真核生物的RNA链的延长。该分子的平面结构部分可以插入至含有连续G=C 碱基配对的DNA双螺旋内部,改变DNA的结构,从而阻止RNA聚合酶沿着DNA模板的移动。放线菌 素D是发现最早的具有抗癌作用的抗生素,作为化疗药物使用,但也是一种致癌剂。它也是一种常用 的分子生物学研究试剂。另一种有机化合物吖啶(acridine),其结构与放线菌素D的平面结构部分相 似,通过类似的机制抑制RNA合成。 (三】终止阶段 与转录起始需要特殊的启动子序列类似,转录终止也主要是由被转录基因末端的一些特殊的DNA 序列信号所引发,使RNA聚合酶离开DNA模板并释放合成的RNA链。这些引发转录终止的DNA序 列信号就称为终止子(terminator)。原核生物的转录终止机制包括依赖p因子与不依赖p因子两种,前 者需要一种ρ因子蛋白参与,而后者则不需要,这两种终止机制分别使用不同的终止子。 1.依赖p因子的转录终止在依赖p因子的转录终止中,终止子通常是包含有一段富含CA的称 为ut(tho utilization)位点(rut site)的序列。含有rut位点序列的RNA转录产物可招募p因子。p因子 280

第十二章A的生物合成 是由六个相同亚基组成的环状蛋白质,能与离开RNA聚合酶的单链RNA结合。p因子还具有ATP酶 活性和解旋酶活性。因子在特定位点与NA转录产物结合后沿若RNA鞋向下游移动直至转录复合 物。P因子引发转录终止的详细机制目前仍不太清楚。但最新的研究结果提示p因子极有可能是通过 引发RNA聚合酶的构象改变而导致延长复合物停止并进而解体,。因子的解旋酶活性也可使RNA特 从DNA模板单链上分开而得以释放(图12-7A)。 (A)依赖因子的转录终止 (B)不依p因子的转录终止 RNA聚合例 site ATP ADP+P 图127原核生物基因转录终止的机制 2.不依赖p因子的转录终止在不依赖ρ因子的转录终止中,其终止子含有两段特殊的序列元 件:约20p长的反向重复序列和紧随其后的连续的约8个A一T碱基对。这段终止子序列一旦被转录 为RNA单链后,反向重复序列即可通时链内形成碱基互补配对形成茎-环(stm-ooD)结构,或称发夹结 构(hairpin structure)。这种发夹结构的形成可破坏随后的延长复合物中RNA-DNA杂化双链中的A U碱基配对(注意A一U配对是最弱的碱基配对),也可破坏转录产物RNA与RNA聚合酶的相互作 用,从而促进RNA的释放和延长复合物的解体(图12-7B和图12-8)。 二、真核生物转录的基本过程 真核生物与原核生物转录的基本过程和机制大致相同,但更为复杂(表124)。真核生物和原核生 物的RNA聚合酶种类不同,结合模板的特性不一样。原核生物RNA聚合酶可直接结合DNA模板,而 真核生物RNA聚合酶需与辅助因子结合后才结合模板,所以两者的转录起始过程有较大区别。原核生 物没有核膜,其转录和翻译同步进行,而真核生物的转录与翻译分别在细胞核和细胞质中进行,大部分 的真核生物RNA需要经过加工修饰后才能成为成熟RNA 281