
生物大分子波谱学原理 吴季辉 7.3主链三共振序列指认 三维,四维三共振实验对于大蛋 白质的核磁研究具有特别的意义, 因为这些实验提供了蛋白质主链以 及部分侧链原子的明确无误的谱峰 证认手段
7.3主链三共振序列指认 生物大分子波谱学原理 吴季辉 三维,四维三共振实验对于大蛋 白质的核磁研究具有特别的意义, 因为这些实验提供了蛋白质主链以 及部分侧链原子的明确无误的谱峰 证认手段
生物大分子波谐学原理 吴季辉 7.4三共振谱 多肽链的结构特点: 个氨基酸残基的H自旋由一个或多个自旋体系组 成,而同核谱中一个自旋体系必定属于同一个残基 同一个残基及相邻残基的H之间距离可能比较近, 但由于蛋白质空间折叠的缘故,蛋白质一级序列上 相隔甚远的残基的H也可能距离比较近 H-C-H H-C-H H H
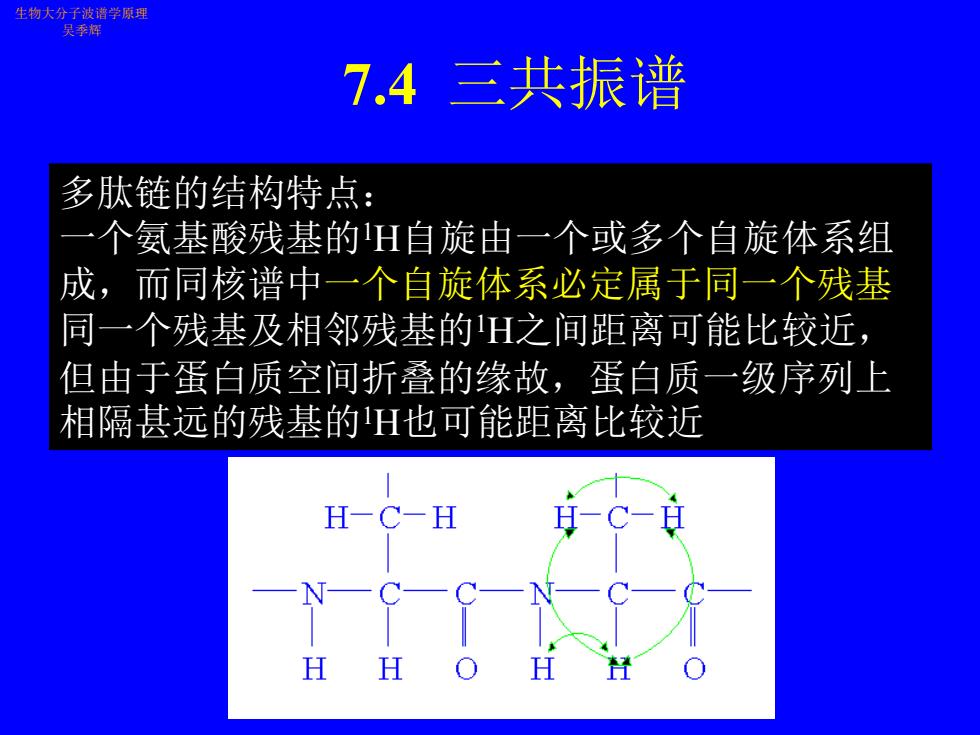
7.4 三共振谱 生物大分子波谱学原理 吴季辉 多肽链的结构特点: 一个氨基酸残基的1H自旋由一个或多个自旋体系组 成,而同核谱中一个自旋体系必定属于同一个残基 同一个残基及相邻残基的1H之间距离可能比较近, 但由于蛋白质空间折叠的缘故,蛋白质一级序列上 相隔甚远的残基的1H也可能距离比较近

生物大分子波谱学原理 吴季辉 7.4三共振谱 同核H谱提供的信息: COSY 提供同碳偶合(geminal coupling)及邻 位偶合(vicinal coupling)的H信息 TOCSY 提供同一自旋体系的H信息 因此COSY类型实验及TOCSY只能给出同 残基内的H的连接信息 NOESY提供空间距离近的H信息,通常有 NOE的H处于同一残基或相邻残基,但也有一部 分(可能达10%)属于相隔一个残基以上的H之 间的NOE
7.4 三共振谱 生物大分子波谱学原理 吴季辉 同核1H谱提供的信息: COSY 提供同碳偶合(geminal coupling)及邻 位偶合(vicinal coupling)的1H信息 TOCSY 提供同一自旋体系的1H信息 因此COSY类型实验及TOCSY只能给出同 一残基内的1H的连接信息 NOESY 提供空间距离近的1H信息,通常有 NOE的1H处于同一残基或相邻残基,但也有一部 分(可能达10%)属于相隔一个残基以上的1H之 间的NOE

生物大分子波谐学原理 吴季辉 7.4三共振谱 在同核H谱基础上的谱峰证认手段: 由同核相关谱找出谱峰所属的自旋体系,判别可 能的氨基酸类型 由NOESY结合已知的蛋白质序列进一步确定氨 基酸类型并确定其在蛋白质序列上的位置 由于序列证认依靠NOESY,有错误证认的可能, 通常需要在获得初步结构的过程中反复check NOE,纠正可能的误认,因而实际上序列证认与 结构计算不可完全分开
7.4 三共振谱 生物大分子波谱学原理 吴季辉 在同核1H谱基础上的谱峰证认手段: 由同核相关谱找出谱峰所属的自旋体系,判别可 能的氨基酸类型 由NOESY结合已知的蛋白质序列进一步确定氨 基酸类型并确定其在蛋白质序列上的位置 由于序列证认依靠NOESY,有错误证认的可能, 通常需要在获得初步结构的过程中反复check NOE,纠正可能的误认,因而实际上序列证认与 结构计算不可完全分开

生物大分子波谱学原理 吴季辉 7.4三共振谱 13C,15N双标记蛋白质的异核三共振谱利用单 键和两键J偶合建立主链原子和部分侧链原子间 的关联,既可提供残基内的,也可提供相邻残基 的HN,15N,1H,13Ca,13CO等原子间的关联信息, 即完成序列证认,由于主要信息来自J偶合,在 结构计算前即可完成大部分,至少主链部分的所 有原子的谱峰证认。既提高证认的可靠性,又加 快证认的速度。而且从获得的主链原子的化学位 移还可判定残基的类型以及所处的二级结构类型
7.4 三共振谱 生物大分子波谱学原理 吴季辉 13C, 15N双标记蛋白质的异核三共振谱利用单 键和两键J偶合建立主链原子和部分侧链原子间 的关联,既可提供残基内的,也可提供相邻残基 的1HN , 15N,1H , 13C , 13CO等原子间的关联信息, 即完成序列证认,由于主要信息来自J偶合,在 结构计算前即可完成大部分,至少主链部分的所 有原子的谱峰证认。既提高证认的可靠性,又加 快证认的速度。而且从获得的主链原子的化学位 移还可判定残基的类型以及所处的二级结构类型