
生物大分子波谱学原理 吴季辉 异核编辑谱 般异核编辑谱由同核NOESY.或TOCSY 同HSQC或HMQC串接成,提供的信息 类似同核谱,但是谱峰在与H核相关的 13C或15N核的化学位移上展开以解决同 核谱重叠的问题。其中异核编辑的 NOESY谱是最后结构计算所需的NOE的 主要来源
异核编辑谱 生物大分子波谱学原理 吴季辉 一般异核编辑谱由同核NOESY或TOCSY 同HSQC或HMQC串接成,提供的信息 类似同核谱,但是谱峰在与1H核相关的 13C或15N核的化学位移上展开以解决同 核谱重叠的问题。其中异核编辑的 NOESY谱是最后结构计算所需的NOE的 主要来源

生物大分子波谱学原理 吴季辉 7.2异核编辑谱 7.2.1 3D NOESY-HSQC ①2 1 t/2 3 decouple 上图是基本的NOESY-HSQC三维谱脉冲序列,适合于单标记的蛋白质样品。所用相 循环为1=2(x),2(-x),2=4(x),4(-x),3=X,-x;rcf=x,-x,-x,x,-xx,x,-x,spin-lock的时间1-2ms。 直至a点,序列是同核NOESY谱序列,由于现在是标记样品,故t维演化时要加异 核180度脉冲实现去偶,也可用异核宽带去偶。NOESY部分的最后一个90度H脉冲相 当于HSQC中第一个H90度脉冲,剩余部分同H$QC序列。其中的自旋锁定脉冲用于抑 制水峰。也可利用梯度场脉冲抑制水峰。在实际使用时,若是用于1N标记样品,由于重 要信息是HNH,通常不用预饱和以免H被衰减。这时为了提高水峰抑制效果,最后 个回波的1H180度脉冲常用3-9-19形式的WATERGATE
7.2 异核编辑谱 生物大分子波谱学原理 吴季辉

生物大分子波谱学原理 吴季辉 7.2异核编辑谱 其中6个H脉冲的宽度 依次为0.231*90, 0.692*90,1.462*90, 1.462*90,0.692*90 0.231*90,间隔t取300微 秒左右(500兆上),而 1 5x+2x1= ,这种 2Jg 形式的WATERGATE抑 制水峰效果较好。6个1H 脉冲的等价效果在照射 点为0度,在NH上为180度,因此激发曲线在H附近有很大衰减,但此时由于15N编 辑的缘故,观察的信号只能是NH,故不会导致什么问题。若是用于13C标记样品,则不 能用这种WATERGATE,但一般13C标记样品溶于重水,故水峰抑制不成问题。不过对 于全标记的13C样品,需考虑13CO的去偶,以及C-C间的偶合
7.2 异核编辑谱 生物大分子波谱学原理 吴季辉
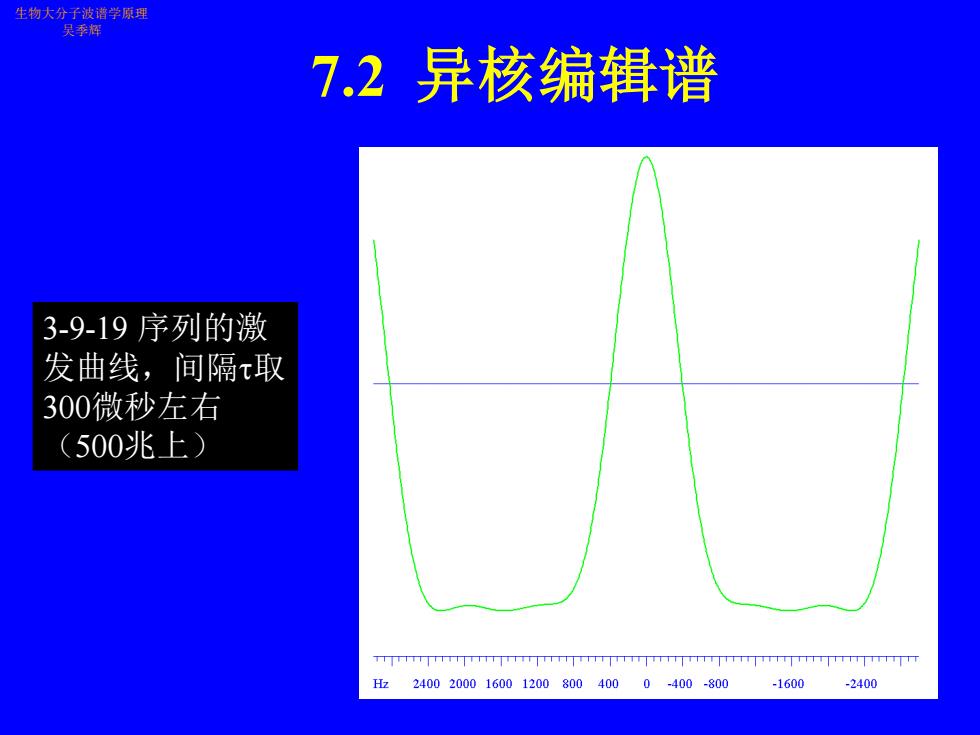
生物大分子波谱学原理 吴季辉 7.2异核编辑谱 3-9-19序列的激 发曲线,间隔取 300微秒左右 (500兆上) npinpinpinnpnnpinnpn 224002000.160012008004000-400-800-1600 -2400
7.2 异核编辑谱 生物大分子波谱学原理 吴季辉 3-9-19 序列的激 发曲线,间隔取 300微秒左右 (500兆上)

生物大分子波谐学原理 吴季辉 7.2异核编辑谱 整个序列的信号传递过程可以简化成: K,(传)0→1,s→S,(5)s→1,(馬) 作三维谱同二维谱的一个重要区别在于数据点的选择,由于实验时间的限制, 间接维点数不能太多,这个谱的间接维点数可取128×32复数点。15N维由于N核数 目不多,而且一般化学位移重叠不很严重,取几十点即可,H维应该取得大一些, 当然要考虑到实验时间的限制。由于NOESY谱的重要性,信噪比要求比较高,经常 要采样3一4天甚至更长。 序列中的HSQC部分可用其他异核相关谱代替,如HMQC,尽管二维谱HMQC 有分辨率低的缺点,在三维谱中表现不是太差,因为三维谱分辨率的主要限制在数 据点数的大小上
7.2 异核编辑谱 生物大分子波谱学原理 吴季辉