
活化G蛋白与其他蛋白质结合传递信号 GTP结合后,G蛋白与By亚基相互作用的表面发生变化,G蛋白对By二聚体的亲和性 大大降低。但是G蛋白所采用的新结构与其他蛋白质有亲和性。在阝-AR途径,与G蛋白 发生结合的新蛋白是腺苷酸环化酶(adenylate cyclase)。腺苷酸环化酶是含有l2个跨膜螺旋 的膜蛋白,能够将ATP转化成CAMP。环化酶两个大的细胞质结构域形成酶的催化区。GCs 与腺苷酸环化酶相互作用有利于环化酶的催化活性,促进cAMP的产出(图14.7)。实际上, 参与B-AR途径的G蛋白亚基是GS(s表示具有促进作用)。肾上腺素与细胞表面接合的 净结果是增加了胞内cAMP合成速率。腺苷酸环化酶催化合成的CAMP为信号放大提供了 第二水平,因为每个腺苷酸环化酶能够将很多ATP分子转化成cAMP。 CAMP活化蛋白激酶A,后者催化很多目标蛋白磷酸化 我们继续跟踪信号传导途径的信息流动。CAMP浓度的增加能影响不同的细胞过程。在 肌肉组织的肾上腺素途径,CAMP刺激肌肉细胞合成ATP以利于肌肉收缩。在其他细胞, CAMP促进储存燃料的降解、增加胃粘膜分泌酸,导致黑素颗粒的分散,消除血小板凝聚, 诱导氯离子通道的开放。cAMP如何影响如此之多的细胞过程?实际上,真核细胞cAMP 的大多数效应是由于一种蛋白激酶的活化所致。这个蛋白质是蛋白激酶A(PKA)。 如同早先介绍的,PKA有两个调节亚基(R)和两个催化亚基(C)。没有cAMP,R2C2 复合物没有催化活性。CAMP与调节亚基结合,就能释放出具有催化亚基。催化亚基催化自 身磷酸化。活化的PKA能够将很多目标蛋白特定位点的丝氨酸和酸酸磷酸化。例如,PKA 磷酸化两种酶(这两种酶促经糖原降解,抑制糖原合成)(21.3节)。PKA磷酸化一些转录 因子(这些转录因子称为cAMP应答因子结合蛋白,即CREB,cAMP-response element binding protein),从而刺激特定基因的表达。PKA的这种活性显示信号转导途径能够将胞外信息传 递到细胞核,改变基因的转录。 图14.8总结了肾上腺素启动的信号转导途径。 Epinephrine B-Adrenergic receptor Binding Activated receptor GTP for GDP exchange Amplification Activated G protein Activated adenylate cyclase Enzymatic reaction Amplification Increased [cAMP] Activated protein kinase A and other effectors Figure 14-8 2007 W.H.Freeman and Compary 图14.8肾上腺素信号传递途径。肾上腺素与肾上腺素受体结合,启动信号传导途径。每步 反应的特性在箭头左侧的黑色文字标出。箭头右侧绿色文字标出能进行信号放大的步骤
活化 G 蛋白与其他蛋白质结合传递信号 GTP 结合后,G 蛋白与 亚基相互作用的表面发生变化,G 蛋白对 二聚体的亲和性 大大降低。但是 G 蛋白所采用的新结构与其他蛋白质有亲和性。在 AR 途径,与 G 蛋白 发生结合的新蛋白是腺苷酸环化酶(adenylate cyclase)。腺苷酸环化酶是含有 12 个跨膜螺旋 的膜蛋白,能够将 ATP 转化成 cAMP。环化酶两个大的细胞质结构域形成酶的催化区。Gs 与腺苷酸环化酶相互作用有利于环化酶的催化活性,促进 cAMP 的产出(图 14.7)。实际上, 参与 -AR 途径的 G 蛋白亚基是 Gs(s 表示具有促进作用)。肾上腺素与细胞表面接合的 净结果是增加了胞内 cAMP 合成速率。腺苷酸环化酶催化合成的 cAMP 为信号放大提供了 第二水平,因为每个腺苷酸环化酶能够将很多 ATP 分子转化成 cAMP。 cAMP 活化蛋白激酶 A,后者催化很多目标蛋白磷酸化 我们继续跟踪信号传导途径的信息流动。cAMP 浓度的增加能影响不同的细胞过程。在 肌肉组织的肾上腺素途径,cAMP 刺激肌肉细胞合成 ATP 以利于肌肉收缩。在其他细胞, cAMP 促进储存燃料的降解、增加胃粘膜分泌酸,导致黑素颗粒的分散,消除血小板凝聚, 诱导氯离子通道的开放。cAMP 如何影响如此之多的细胞过程?实际上,真核细胞 cAMP 的大多数效应是由于一种蛋白激酶的活化所致。这个蛋白质是蛋白激酶 A (PKA)。 如同早先介绍的,PKA 有两个调节亚基(R)和两个催化亚基(C)。没有 cAMP, R2C2 复合物没有催化活性。cAMP 与调节亚基结合,就能释放出具有催化亚基。催化亚基催化自 身磷酸化。活化的 PKA 能够将很多目标蛋白特定位点的丝氨酸和酸酸磷酸化。例如,PKA 磷酸化两种酶(这两种酶促经糖原降解,抑制糖原合成)(21.3 节)。PKA 磷酸化一些转录 因子(这些转录因子称为 cAMP 应答因子结合蛋白,即 CREB, cAMP-response element binding protein),从而刺激特定基因的表达。PKA 的这种活性显示信号转导途径能够将胞外信息传 递到细胞核,改变基因的转录。 图 14.8 总结了肾上腺素启动的信号转导途径。 图 14.8 肾上腺素信号传递途径。肾上腺素与肾上腺素受体结合,启动信号传导途径。每步 反应的特性在箭头左侧的黑色文字标出。箭头右侧绿色文字标出能进行信号放大的步骤
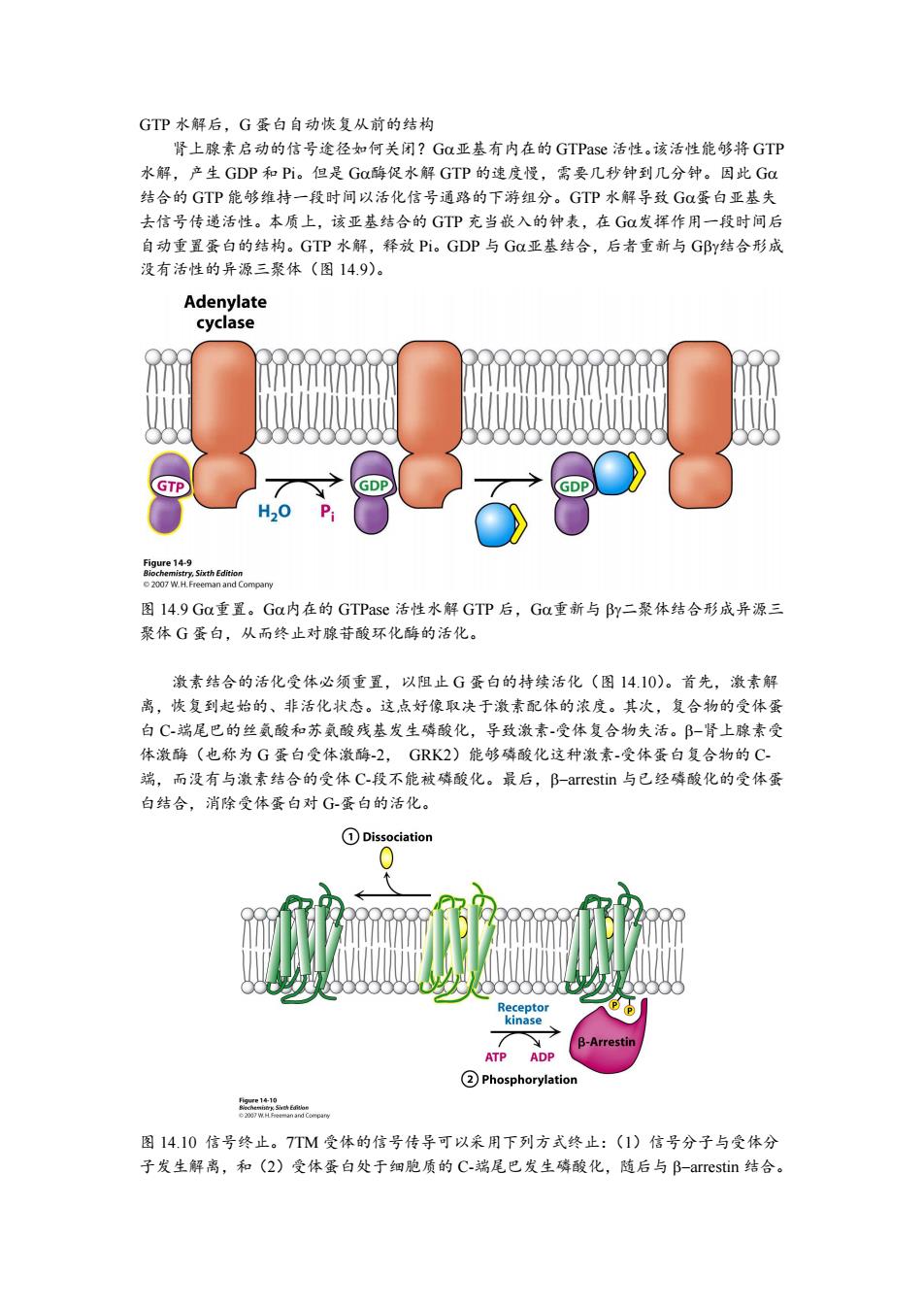
GTP水解后,G蛋白自动恢复从前的结构 肾上腺素启动的信号途径如何关闭?G0亚基有内在的GTPase活性。该活性能够将GTP 水解,产生GDP和Pi。但是Ga酶促水解GTP的速度慢,需要几秒钟到几分钟。因此GC 结合的GTP能够维持一段时间以活化信号通路的下游组分。GTP水解导致G蛋白亚基失 去信号传递活性。本质上,该亚基结合的GTP充当嵌入的钟表,在G发挥作用一段时间后 自动重置蛋白的结构。GTP水解,释放Pi。GDP与GC亚基结合,后者重新与GBy结合形成 没有活性的异源三聚体(图14.9)。 Adenylate cyclase GTP GDP H2O Pi Oee135hEn 2007 W.H.Freeman and Compary 图l4.9Ga重置。Ga内在的GTPase活性水解GTP后,Ga重新与By二聚体结合形成异源三 聚体G蛋白,从而终止对腺苷酸环化酶的活化。 激素结合的活化受体必须重置,以阻止G蛋白的持续活化(图14.10)。首先,激素解 离,恢复到起始的、非活化状态。这点好像取决于激素配体的浓度。其次,复合物的受体蛋 白C-端尾巴的丝氨酸和苏氨酸残基发生磷酸化,导致激素-受体复合物失活。B-肾上腺素受 体激酶(也称为G蛋白受体激酶-2,GK2)能够磷酸化这种激素-受体蛋白复合物的C 端,而没有与激素结合的受体C-段不能被磷酸化。最后,B-arrestin与已经磷酸化的受体蛋 白结合,消除受体蛋白对G-蛋白的活化。 ①Dissociation Receptor kinase y B-Arrestin ATP ADP ②Phosphorylation 图14.10信号终止。7TM受体的信号传导可以采用下列方式终止:(1)信号分子与受体分 子发生解离,和(2)受体蛋白处于细胞质的C-端尾巴发生磷酸化,随后与B-arrestin结合
GTP 水解后,G 蛋白自动恢复从前的结构 肾上腺素启动的信号途径如何关闭?G亚基有内在的 GTPase 活性。该活性能够将 GTP 水解,产生 GDP 和 Pi。但是 G酶促水解 GTP 的速度慢,需要几秒钟到几分钟。因此 G 结合的 GTP 能够维持一段时间以活化信号通路的下游组分。GTP 水解导致 G蛋白亚基失 去信号传递活性。本质上,该亚基结合的 GTP 充当嵌入的钟表,在 G发挥作用一段时间后 自动重置蛋白的结构。GTP 水解,释放 Pi。GDP 与 G亚基结合,后者重新与 G结合形成 没有活性的异源三聚体(图 14.9)。 图 14.9 G重置。G内在的 GTPase 活性水解 GTP 后,G重新与 二聚体结合形成异源三 聚体 G 蛋白,从而终止对腺苷酸环化酶的活化。 激素结合的活化受体必须重置,以阻止 G 蛋白的持续活化(图 14.10)。首先,激素解 离,恢复到起始的、非活化状态。这点好像取决于激素配体的浓度。其次,复合物的受体蛋 白 C-端尾巴的丝氨酸和苏氨酸残基发生磷酸化,导致激素-受体复合物失活。肾上腺素受 体激酶(也称为 G 蛋白受体激酶-2, GRK2)能够磷酸化这种激素-受体蛋白复合物的 C- 端,而没有与激素结合的受体 C-段不能被磷酸化。最后,arrestin 与已经磷酸化的受体蛋 白结合,消除受体蛋白对 G-蛋白的活化。 图 14.10 信号终止。7TM 受体的信号传导可以采用下列方式终止:(1)信号分子与受体分 子发生解离,和(2)受体蛋白处于细胞质的 C-端尾巴发生磷酸化,随后与 arrestin 结合

一些7TM受体能够活化磷酸肌醇级联反应 现在看另一个常见的第二信使级联反应。很多激素的受体也是TTM受体。磷酸肌醇级 联反应与cAMP级联反应一样,将细胞外的信息转化成细胞内信号分子。磷脂酰肌醇-4,5- 二磷酸(PP2)是细胞膜上的一种磷脂分子。信号转导途径导致该磷脂分子水解产生第二信 使。血管紧张素Ⅱ受体与控制血压的激素结合,启动的信号途径就产生这样的第二信使就 是这样的实例。 每类TTM受体所结合的G蛋白不同。肾上腺素与受体结合将G蛋白活化成G0S,血管 紧张素Ⅱ与受体结合所活化的G蛋白叫G0q。与GTP结合时,GCq能结合并活化B类磷脂 酶C。后者催化PIP2断裂形成两种第二信使,即肌酸1,4,5-三磷酸(IP3)和二酯酰甘油(DAG: 图14.11)。 Phospholipase C Phosphatidylinositol 4,5-bisphosphate(PIP,) OH OPO o, Diacylglycerol (DAG) Inositol 1,4,5-trisphosphate(IP3) Rigure 14-11 图14.11磷脂酶C酶促反应。磷脂酶C断裂膜磷脂成分磷脂酰肌醇-4,5-二磷酸,产生二酯 酰甘油(仍在膜上)和肌醇-1,4,5-三磷酸(水溶分子,在细胞质扩散)。 P3是可溶性分子,从生物膜离开进入进入细胞质溶液扩散。这个第二信使作用于C+ 库内质网膜的转移器(如Ca2+-ATPase),导致Ca2+在胞内迅速释放。P3与内质网Ca2+通道 的特异结合,通道开放使Ca+从内质网流入细胞质。Ca+本身也是第二信使:它能结合很多 蛋白质,包括普遍存在的信号传递蛋白即钙调蛋白(calmodulin)和酶(如蛋白激酶C)。采用 这些方式,C+浓度的提升能启动很多生理过程,包括平滑肌收缩、糖原分解、和囊泡释放。 DAG仍然停留在细胞膜上,但是它能够活化蛋白激酶C。蛋白激酶C能够使很多目标 蛋白的丝氨酸和苏氨酸发生磷酸化。蛋白激酶C的DAG结合域需要Ca2+才能与DAG结合。 二酰基甘油和IP3联合作用:IP3增加Ca2+浓度,Ca2+浓度提高有助于蛋白激酶C活化。图 14.12总结了磷酸肌醇级联反应。由于IP3和DAG已经通过磷酸化或其他生物过程转化成 信息,这两种第二信使的作用时间很短
一些 7TM 受体能够活化磷酸肌醇级联反应 现在看另一个常见的第二信使级联反应。很多激素的受体也是 7TM 受体。磷酸肌醇级 联反应与 cAMP 级联反应一样,将细胞外的信息转化成细胞内信号分子。磷脂酰肌醇-4,5- 二磷酸(PIP2)是细胞膜上的一种磷脂分子。信号转导途径导致该磷脂分子水解产生第二信 使。血管紧张素-II 受体与控制血压的激素结合,启动的信号途径就产生这样的第二信使就 是这样的实例。 每类 7TM 受体所结合的 G 蛋白不同。肾上腺素与受体结合将 G 蛋白活化成 Gs,血管 紧张素 II 与受体结合所活化的 G 蛋白叫 Gq。与 GTP 结合时,Gq 能结合并活化 类磷脂 酶 C。后者催化 PIP2 断裂形成两种第二信使,即肌酸 1,4,5-三磷酸(IP3)和二酯酰甘油(DAG; 图 14.11)。 图 14.11 磷脂酶 C 酶促反应。磷脂酶 C 断裂膜磷脂成分磷脂酰肌醇-4,5-二磷酸,产生二酯 酰甘油(仍在膜上)和肌醇-1,4,5-三磷酸(水溶分子,在细胞质扩散)。 IP3 是可溶性分子,从生物膜离开进入进入细胞质溶液扩散。这个第二信使作用于 Ca 2+ 库内质网膜的转移器(如 Ca 2+ -ATPase),导致 Ca 2+在胞内迅速释放。IP3 与内质网 Ca 2+通道 的特异结合,通道开放使 Ca 2+从内质网流入细胞质。Ca 2+本身也是第二信使:它能结合很多 蛋白质,包括普遍存在的信号传递蛋白即钙调蛋白(calmodulin)和酶(如蛋白激酶 C)。采用 这些方式,Ca 2+浓度的提升能启动很多生理过程,包括平滑肌收缩、糖原分解、和囊泡释放。 DAG 仍然停留在细胞膜上,但是它能够活化蛋白激酶 C。蛋白激酶 C 能够使很多目标 蛋白的丝氨酸和苏氨酸发生磷酸化。蛋白激酶 C 的 DAG 结合域需要 Ca 2+才能与 DAG 结合。 二酰基甘油和 IP3 联合作用:IP3 增加 Ca 2+浓度,Ca 2+浓度提高有助于蛋白激酶 C 活化。图 14.12 总结了磷酸肌醇级联反应。由于 IP3 和 DAG 已经通过磷酸化或其他生物过程转化成 信息,这两种第二信使的作用时间很短

Diacylglycerol(DAG) Cell membrane DAG PIP2 Phospholipase C cleavage Calcium ion Protein kinase C Cytoplasm receptor ER membrane Calciumo 00 ion 0 0 00 2007 W.H.Freeman and Company 图14.12磷脂酰肌醇级联反应。磷脂酰肌醇4,5-二磷酸(PIP2)断裂形成二脂酰甘油(DAG) 和肌醇1,4,5-三磷酸(IP3),导致Ca+释放(P3受体的离子通道开放)、活化蛋白激酶C(因 蛋白激酶C与膜上游离的DAG结合)。C+与蛋白激酶C结合有助于蛋白激酶C的活化。 Ca2+是广泛使用的第二信使 除了磷脂酰肌醇级联途径外,C+参与了很多信号传递过程。这个离子有几种特性是指 能够在细胞内广泛充当第二信使。首先,胞内Ca+浓度波动较大。在稳定状态,胞内C+ 浓度水平低,阻止生物分子的羧基或磷酸基团与Ca+结合形成沉淀。将Ca+外排,维持胞 内低浓度Ca2+。由于有这种外排作用,细胞质的Ca+浓度维持在100nM左右,比细胞外的 Ca2+浓度低几个数量级。由于稳态时Ca+浓度低,易于感知信号传递造成的Ca+浓度上升。 第二个特性是它与蛋白质结合紧密并能诱导结合蛋白发生构象变化(图14.13)。Ca2+ 与带负电荷的氧原子(由谷氨酸和天冬氨酸侧链提供)和不带电荷的氧原子(主链我基及谷 氨酰胺和天冬酰胺提供)结合牢。C+与六至八个氧原子形成配位键,将蛋白质不同区域交 联在一起,诱导蛋白质发生构象变化。 探测胞内Ca2+浓度变化(甚至是进行即时检测)方法的建立极大地促进了人们对Ca+ 在胞内生物作用的认识。这个方法有赖于人们设计的染料分子如Fura-2。Fura-2能结合Ca+, 与Ca2+结合后,Fura-2的荧光性质发生改变
图 14.12 磷脂酰肌醇级联反应。磷脂酰肌醇 4,5 - 二磷酸(PIP2)断裂形成二脂酰甘油(DAG) 和肌醇 1,4,5-三磷酸(IP3),导致 Ca 2+释放(IP3受体的离子通道开放)、活化蛋白激酶 C(因 蛋白激酶 C 与膜上游离的 DAG 结合)。Ca 2+与蛋白激酶 C 结合有助于蛋白激酶 C 的活化。 Ca 2+是广泛使用的第二信使 除了磷脂酰肌醇级联途径外,Ca 2+参与了很多信号传递过程。这个离子有几种特性是指 能够在细胞内广泛充当第二信使。首先,胞内 Ca 2+浓度波动较大。在稳定状态,胞内 Ca 2+ 浓度水平低,阻止生物分子的羧基或磷酸基团与 Ca 2+结合形成沉淀。将 Ca 2+外排,维持胞 内低浓度 Ca 2+。由于有这种外排作用,细胞质的 Ca 2+浓度维持在 100 nM 左右,比细胞外的 Ca 2+浓度低几个数量级。由于稳态时 Ca 2+浓度低,易于感知信号传递造成的 Ca 2+浓度上升。 第二个特性是它与蛋白质结合紧密并能诱导结合蛋白发生构象变化(图 14.13)。Ca 2+ 与带负电荷的氧原子(由谷氨酸和天冬氨酸侧链提供)和不带电荷的氧原子(主链羰基及谷 氨酰胺和天冬酰胺提供)结合牢。Ca 2+与六至八个氧原子形成配位键,将蛋白质不同区域交 联在一起,诱导蛋白质发生构象变化。 探测胞内 Ca 2+浓度变化(甚至是进行即时检测)方法的建立极大地促进了人们对 Ca 2+ 在胞内生物作用的认识。这个方法有赖于人们设计的染料分子如 Fura-2。Fura-2 能结合 Ca 2+, 与 Ca 2+结合后,Fura-2 的荧光性质发生改变