
第十三章蛋白质的生物合成 位结合氨基酰RNA,称氨基酰位(aminiacyl site):P位结合肽酰-RNA,称肽酰位(peptidyl site):E位是 排出位(exit site),由此释放已经卸载了氨基酸的RNA。真核生物的核糖体上没有E位,空载的tRNA 直接从P位脱落。 四、蛋白质合成体系的其他组分 蛋白质生物合成需要由ATP和GTP提任能最需要M:肽稀转移德(成称转肽酶),氨甚酰RNA 合成酶等多种酶参与反应,从起始、延长到终止的各阶段还需要多种核糖体以外的其他蛋白因子(表 13-2、表13-3)。这些因子有:①①起始因子(initiation factor,F),原核生物(prokaryote)和直核生物(eu karyote)的起始因子分别用和eF表示:②延长因子(nion factor,EF),原核生物与真核生物的延 长因子分别用EF和eEF表示;③释放因子(release factor,.RF)又称终止因子(termination factor),原核生 物与真核生物的释放因子分别用RF和eRF表示 表132原核生物肽链合成所需要的蛋白因子 种类 生物学功能 起始因子 占据核糖体A位,防止A位结合其他RNA IF-2 促进Me-RNA与小亚基结合 F-3 促进大、小亚基分离;提高P位结合Met-tRNA的敏感性 延长因子 EF-Tu 促进氨基酰-RNA进入A位,结合并分解GTP EF-T EFTu的调节亚基 EF-G 有转位酶活性,促进肽酰-RNA由A位移至P位:促进RNA卸载与释放 释放因子 RF-1 特异识别终止密码子UAA,UAG:诱导肽酰转移酶转变为酯酶 RF-2 特异识别终止密码子UAA,UGA:诱导肽酰转移酶转变为酯 RF-3 具有GTP鹂活性,介导RF1及RF2与核糖体的相互作用 表13.3直核生物肽皓合成所需要的蛋白因子 种类 生物学功能 起始因子 IE-I 多功能因子,参与翻译的多个步骤 elF-2 促进Me-RNA,"“与小亚基结合 IF.2B 结合小亚基,促进大、小亚基分离 elF-3 结合小亚基,促进大小亚基分离:介导F-4FmRNA与小亚基结合 elF-4A F-4F复合物成分:有RNA解螺旋酶活性,解除mRNA5'-端的发夹结构,使其与小亚基结合 elF-4B 结合mRNA.协助mRNA扫描定位起始密码子AUG eIF-4E F.4F复合物成分,识别结合mRNA的5'-端的帽结构 elF-40 cF-4F复合物成分,结合F4E,eF3和PAB elF.5 促进各种起始因子从小亚基解离 elF-6 促进大,小亚基分离 延长因子 eEFl-a 促进氨基酰RNA进人A位;结合分解GTP,相当于EFT eEFI-By 调节亚基,相当于EF-T EF-2 有转位酶活性,促进肽酰-RNA由A位移至P位:促进RNA卸载与释放,相当于EFG 释放因子 eRF 识别所有终止密码子,具有原核生物各类RF的功能 3

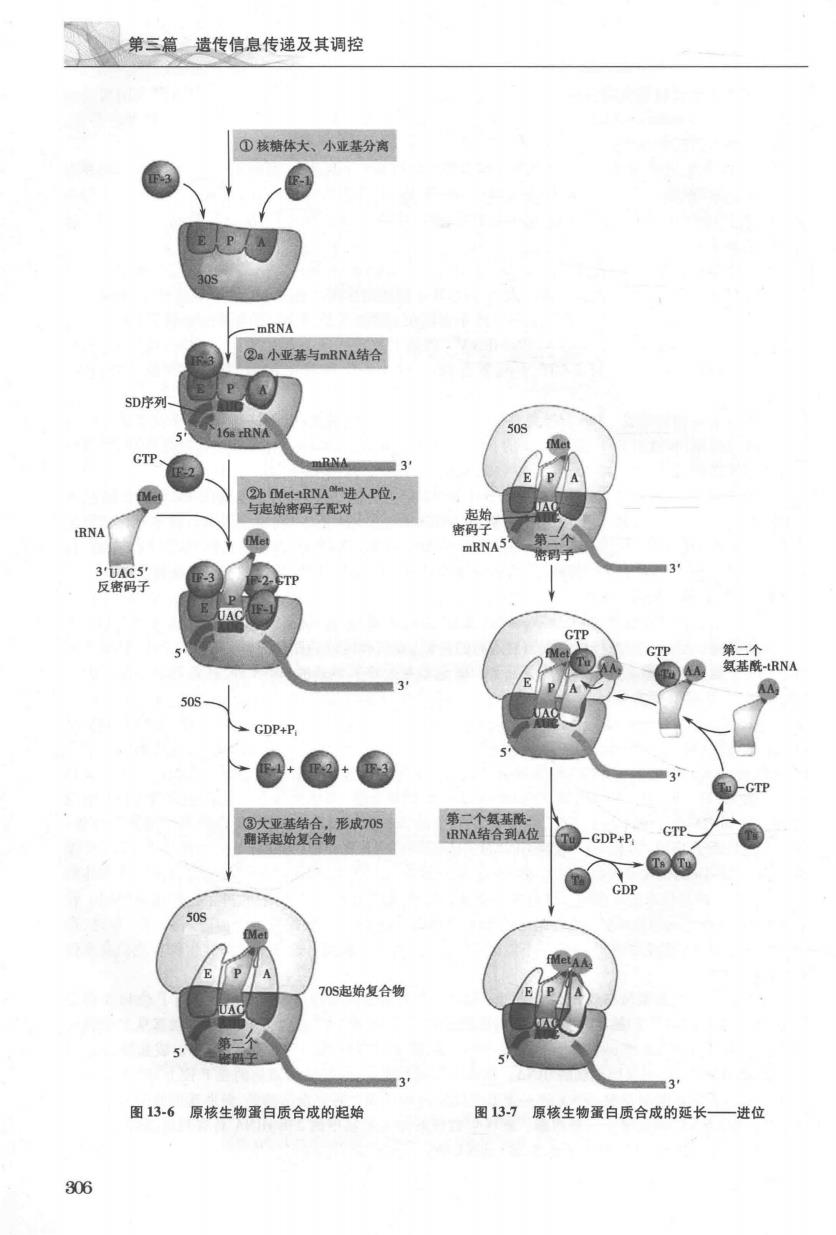
第三篇遗传信息传递及其调控 第二节蛋白质生物合成过程 蛋白质生物合成的过程相当复杂,从氨基端(N-端)向骏基端(C-端)进行,大致可分为以下几个阶 段:①氨基酸活化为氨基酰-RNA;②活化氨基酸转运至核糖体;③活化氨基酸在核糖体上进行缩合反应 合成肽链,此阶段可分为肽链合成的起始、肽链的延长和肽链合成的终止与释放。前两个阶段是蛋白质 生物合成的准备阶段(详见本章第一节),第三个阶段则是蛋白质生物合成的中心环节 一、原核生物蛋白质生物合成过程 原核生物蛋白质生物合成与真核生物有许多相似之处。以氨基酰-RNA形式存在的活化氨基酸 在核糖体上缩合成肽,整个过程划分为起始、延长和终止三个阶段。 (一)翻译起始阶段 蛋白质合成的起始并不是从mRNA的S'端第一个核苷酸开始的。许多原核生物的mRNA分子往 往是多顺反子(polyeistron),即同一mRNA编码相关的多条肽链。在翻译时,每个多肽链都有各自的起 始密码子与终止密码子,决定其合成的起始和终止位置。那么原核细胞中的核糖体是如何对mRNA分 子内如此众多的起始密码子AUG进行识别?在20世纪70年代初期,澳大利亚学者John Shine和Lynn Dalgarno发现,在原核生物mRNA上起始密码子AUG上游的5'端大约10个核苷酸处,存在一段由4~9 个核苷酸组成的富含嘌吟碱基的序列,命名为Shine-Dalgarno序列(SD序列)(图135A)。SD序列正好 与核糖体30S小亚基内部的16SRNA3'端的一段富含密啶碱基的序列互补识别,从而使mRNA和小亚 基结合,因此SD序列又称为核糖体结合位点(ribosomal binding site,RBS)(图135B)。另外,紧邻SD 序列上游的一小段核苷酸又可被核糖体小亚基蛋白印S-1辨认结合,从而协助RNA的起始密码子 AUG对准核糖体的P位。因此,原核生物通过以下两种相互作用确定蛋白质合成的起始部位:①mRNA 的5'端SD序列与16 S rRNA3'端序列的配对:②mRNA上起始密码子AUG与Mt-tRNA=的反密码子 相互识别。细菌毒素colecin E3可通过核酸酶的活性特异性地在6 SrRNA3'端切下50个碱基左右的 片段,使16SRNA失去与mRNA的SD序列互补的部分,由此抑制了细菌蛋白质的合成。由于原核生 物和真核生物在蛋白质合成起始机制上的差异,colecin E3并不影响真核生物核糖体的功能 phage cro (与1s部分序列E配对)(与高对) A 16 S rRNA的3端 BRNA (SGAUUCCUACGAGCUUUGACCUAUGCGACCUUUUACU) B 135原核生物mRNA的SD序列 A五种mRNA的SD序列;BSD序列与I6SRNA的3'端富含密啶的序列互补 304

第十三童白的生物合 在原核生物蛋白质生物合成的起始阶段,核糖体的大、小亚基,mRNA和Met-iRNA“共同构成翻 译起始复合物(translational initiation complex)。这一过程还需要起始因子(表l3-2)、GTP和Mg2“参与。 起始阶段可分为三个步骤: 1.核糖体大,小亚基的分离F-3首先结合到核糖体30S小亚基上,可能在其与50S大亚基的界面 上,故能促进核糖体大小亚基的解离,使核糖体30S小亚基从不具有活性的70S核糖体释放。F-1与小 亚基的A位结合则能加速这种解离,避免起始氨基酰-RNA与A位的提早结合,同时也有利于F2结 合到小亚基上。 2.30S起始复合物的形成核糖体30S小亚基可与mRNA及Met-RNA分别结合。mRNA与核 糖体小亚基的结合可能是蛋白质合成的限速步骤,F-3起辅助作用。通过mRNA5'端的SD序列与小亚 基中16SRNA3'端的互补序列结合,这样30S小亚基在mRNA上的结合位置正好使小亚基上的P位对 准起始密码子AUG。在F-2参与下,Mt-RNA借助其反密码子与起始密码子配对,进人P位,与 GTP共同形成Met-RNAM-IF-2-GTP中间复合物。此时,A位被F-】占据,不与任何氨基酰-RNA 结合 3.70S复合物的形成30S起始复合物一经形成,F3即行脱落,以便核糖体大,小亚基重新结合 形成70S核糖体,释放出F-1,最后F-2的GTP酶活性被激活,水解GTP释放能量并随之脱落,最终形 成完整的翻译起始复合物(即70S核糖体-Met-tRNA-mRNA)(图13-6) 如前所述,原核生物的核糖体上存在着3个RNA的结合区,分别称为氨基酰位(A位)、肽酰位(P 位)和排出位(E位)。在肽链合成中,新的氨基酰-RNA先进人的是A位,形成肽键后移至P位。但是 在翻译起始复合物装配时,结合起始密码子AUG的Mt-RNA直接结合至核糖体P位,A位空留,且 对应于起始密码子AUG后一个密码子,为新的氨基酰-RNA的进入及肽链延长做好了准备。 (二)翻译延长阶段 翻译起始复合物形成后,核糖体从mRNA的5'-端向3'端移动,依据密码子顺序,从N端开始向C 端合成多肽链。这是一个在核糖体上重复进行的进位、成肽和转位的循环过程,每完成1次,肽链上即 可增加1个氨基酸残基。这一过程除了需要70S起始复合物和氨基酰-RNA外,还需要数种延长因子 (表13-2)以及GTP等参与。 L.进位进位(entrance),又称注册(registration),是指一个氨基酰-tRNA按照mRNA模板的指令 进人并结合到核糖体A位的过程。70S起始复合物中的A位是空闲的,并对应着开放阅读框的第二个 密码子讲人A位的氨基酷tRNA种类即由该密码子决定。出步餐需要GTP Mg2征长因子EFTu和 EFTs(图13-7)。EF-Tu与GTP、氨基酰-RNA结合形成复合物,运达至A位。随后氨基酰RNA的反 密码子与mRNA第二个密码子结合,伴随着GTP分解,EF-Tu以EF-Tu-GDP复合物形式离开核糖体。 另一个延长因子EF-Ts催化GTP置换GDP再生成EF-Tu-GTP复合物,为结合下一个氨基酰-RNA作准 备。因此EF-Tu的作用是促进氨基酰-RNA与A位结合,而EF-Ts则促进EF-Tu的再利用。核糖体对 氨基酷-RNA的讲位有校正作用。肽特生物合成以很高速度讲行,征长阶段的每一时程都有时限。在 此时限内,只有正确的氨基酰RNA能迅速发生反密码子密码子互补配对结合而进入A位。反之,错 误的氨基酰-RNA因反密码子-密码子不能配对结合,而从A位解离。这是维持肽链生物合成的高度保 真性的机制之 2.成肽是指肽酰转移酶(或称转肽酶)催化两个氨基酸间肽键形成的反应。此时P位和A位分 别结合着Met-RNAM和氨基酰-tRNA,在转肽酶催化下,Met-RNAM的甲酰甲硫氨基酰基从P位转移 到A位的氨基酰RNA的a氨基上形成第一个肽键,此步骤需要Mg2和K(图13-8)。成肽后A位结 合二肽酰RNA,P位则保留空载的RNA。从第三个氨基酸开始,转肽酶催化的是P位上RNA所连接 的肽酰基与A位新的氨基酰-RNA的α氨基之间肽键的形成。需要指出的是,转肽酶的化学本质不是 蛋白质,而是RNA,因此属于一种核酶。原核生物核糖体大亚基中的23 S rRNA具有转肽酶的活性,在 真核生物中,该酶的活性位于大亚基的28SRNA中。 305
、第三算通传信息传递及其调控 ①核糖体大、小亚基分离 P mRNA ②小亚基与mRNA结合 sD序列 50S RN 0@© To-GTP To-GDP+P. GTP 0 70S起始复合物 3 ■3 图13-6原核生物蛋白质合成的起始 图137原核生物蛋白质合成的延长—进位 306