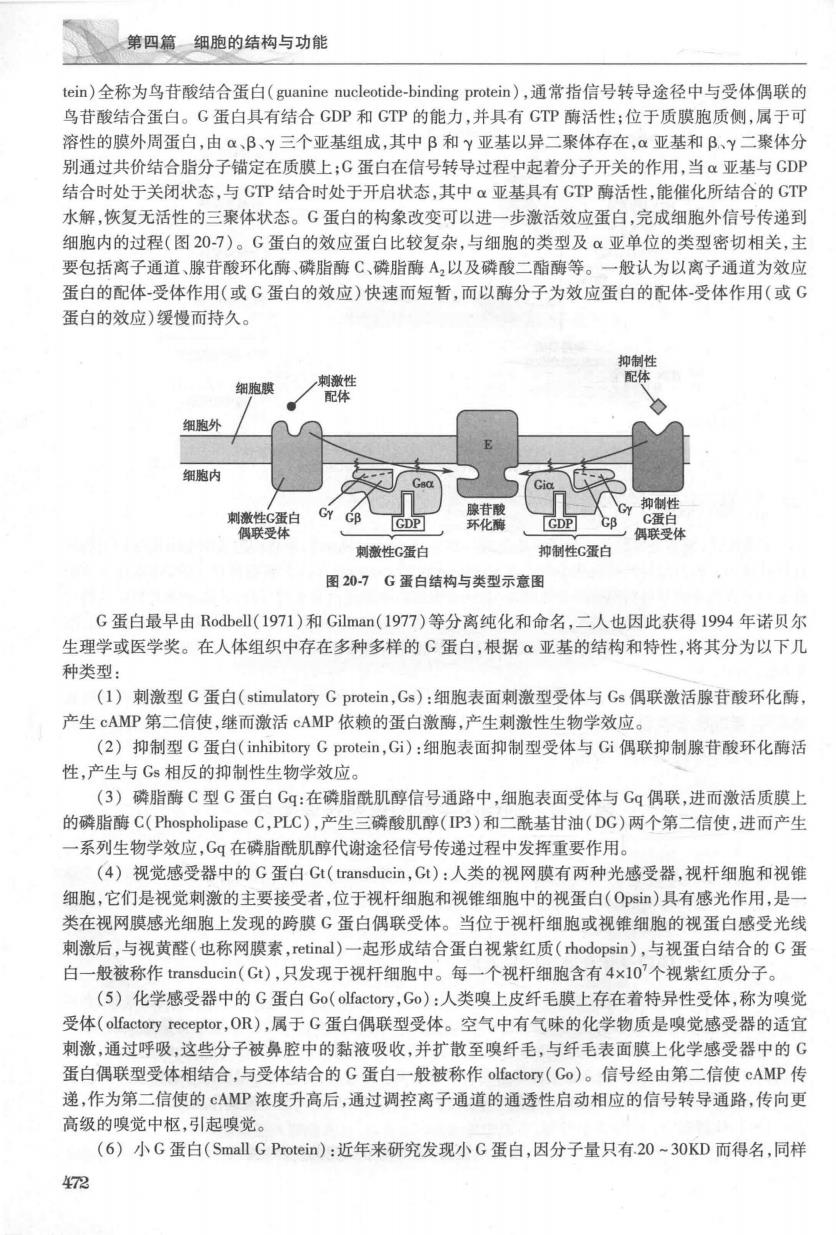
第四篇细胞的结构与功能 tein)全称为鸟苷酸结合蛋白(guanine nucleotide-binding protein),通常指信号转导途径中与受体偶联的 鸟苷酸结合蛋白。G蛋白具有结合GDP和GTP的能力,并具有GTP酶活性;位于质膜胞质侧,属于可 溶性的膜外周蛋白,由a,B、Y三个亚基组成,其中B和y亚基以异二聚体存在,a亚基和B、Y二聚体分 别通过共价结合脂分子锚定在质膜上:G蛋白在信号转导过程中起着分子开关的作用,当α亚基与GDP 结合时处于关闭状态,与GTP结合时处于开启状态,其中α亚基具有GTP酶活性,能催化所结合的GTP 水解,恢复无活性的三聚体状态。G蛋白的构象改变可以进一步激活效应蛋白,完成细胞外信号传递到 细胞内的时程(图207)。G蛋白的数应蛋白出较复杂,与细胞的类型及亚单位的举型密切相关,主 要包括离子通道、腺苷酸环化酶、磷脂酶C、磷脂酶A2以及磷酸二酯酶等。一般认为以离子通道为效应 蛋白的配体受体作用(或G蛋白的效应)快速而短暂,而以酶分子为效应蛋白的配体受体作用(或G 蛋白的效应)缓慢而持久。 细胞眼 CY CB 联受体 刺激性G蛋白 抑制性C蛋白 图20-7G蛋白结构与类型示意图 G蛋白最早由Rodell(1971)和Gilman(1977)等分离纯化和命名,二人也因此获得1994年诺贝尔 生理学或医学奖。在人体组织中存在多种多样的G蛋白,根据α亚基的结构和特性,将其分为以下几 种类型: (1)刺激型G蛋白(stimulatory Gprotein,Gs):细胞表面刺激型受体与Gs偶联激活腺苷酸环化酶, 产生cAMP第二信使,继而激活cAMP依赖的蛋白激酶,产生刺激性生物学效应。 (2)抑制型G蛋白(inhibitory G protein,Gi):细胞表面抑制型受体与Gi偶联抑制腺苷酸环化酶活 性,产生与Gs相反的抑制性生物学效应。 (3)磷脂酶C型G蛋白Gq:在磷脂酰肌醇信号通路中,细胞表面受体与Gg偶联,进而激活质膜上 的磷脂酶C(PhospholipaseC,PLC),产生三磷酸肌醇(IP3)和二酰基甘油(DC)两个第二信使,进而产生 一系列生物学效应,Gq在磷脂酰肌醇代谢途径信号传递过程中发挥重要作用。 (4)视觉感受器中的G蛋白Gt(transducin,Gt):人类的视网膜有两种光感受器,视杆细胞和视锥 细胞,它们是视觉刺激的主要接受者,位于视杆细胞和视雌细胞中的视蛋白(Ops)具有感光作用,是 类在视网膜感光细胞上发现的跨膜G蛋白偶联受体。当位于视杆细胞或视锥细胞的视蛋白感受光线 刺激后,与视黄醛(也称网膜素,retinal)一起形成结合蛋白视紫红质(rhodopsin),与视蛋白结合的G蛋 白一般被称作transducin(Gt),只发现于视杆细胞中。每一个视杆细胞含有4×10'个视紫红质分子。 (5)化学感受器中的G蛋白Go(olfactory,Go):人类嗅上皮纤毛膜上存在着特异性受体,称为嗅觉 受体(olfactory receptor,OR),属于G蛋白偶联型受体。空气中有气味的化学物质是嗅觉感受器的适宜 刺激,通过呼吸,这些分子被鼻腔中的黏液吸收,并扩散至嗅纤毛,与纤毛表面膜上化学感受器中的G 蛋白偶联型受体相结合,与受体结合的G蛋白一般被称作olfactory(Co)。信号经由第二信使cAMP传 递,作为第二信使的cAMP浓度升高后,通过调控离子通道的通透性启动相应的信号转导通路,传向更 高级的嗅觉中枢,引起嗅觉。 (6)小G蛋白(Small G Protein):近年来研究发现小G蛋白,因分子量只有20-30KD而得名,同样 472

第二十章细胞信号转导 具有GP酶活性,在多种细胞反应中具有分子开关作用。第一个被发现的小G蛋白是Ras,它是as基 因的产物。其他的还有Rho、Rab、SEC4、YPT1等。小G蛋白的共同特点是,当结合了GTP时即成为活 化形式,这时可作用于下游分子使之活化,而当GTP水解成为GDP时(自身为GTP酶)则回复到非活化 状态。这一点与Ga类似,但是小G蛋白的分子量明显低于Ga。 Ras蛋白主要参与细胞增殖和信号转导;Rho蛋白对细胞骨架网络的构成发挥调节作用;Rab蛋白 则参与调控细胞内膜交通(membrane traf酯c)。此外,Rho和Rab亚家庭可能分别参与淋巴细胞极化 (polarization)和抗原的提呈。某些信号蛋白通过SH3功能区将酪氨酸激酶途径同一些由小G蛋白所控 制的途径连接起来,如Rh0(与Ras有30%同源性)调节胞浆中微丝上肌动蛋白的聚合或解离,从而影响 细胞形态。这些不同类型的小C蛋白在信号传递过程各种发挥不同的作用。 此外,在细胞中存在着一些专门控制小G蛋白活性的小G蛋白调节因子,有的可以增强小G蛋白 的活性,如鸟苷酸交换因子(guanine nucleotide exchange factor,GEF)和鸟苷酸解离抑制因子(Guanine nucleotide dissociation Inhibitor,GDI),有的可以降低小G蛋白活性,如GTP酶活化蛋白(CTPase activating) (二)G蛋白偶联受体介导的cAMP信号通路 环磷酸腺苷(eyelic AMP,cAMP)是最重要的胞内第二信使,其产生受细胞膜中刺激性受体(Rs)、抑 制性受体(Ri)、刺激型G蛋白(Gs)、抑制性G蛋白(Gi)和腺苷酸环化酶(Adenylate cyclase,AC)五种组 分控制,产生的第二信使cAMP可以被特异的环腺苷酸磷酸二酯酶(cAMP phosphodiesterase)迅速水解 为5-AMP,从而终止信号。 众所周知,肾上腺髓质可分泌肾上腺素和去甲肾上腺素。肾上腺素可以调节糖代谢,促进肝糖原和 肌糖原的分解,增加血糖和血中的乳酸含量。去甲肾上腺素也有类似作用,但作用较弱。肾上腺素由肾 上腺分泌后通过血液输送到肝细胞,即与肝细胞表面上的肾上腺素受体结合。肾上腺素受体属于G蛋 白偶联受体,可分为α及B两个类型。肾上腺素对α、B两型受体均起作用,而去甲肾上腺素主要对α 型受体起作用 一旦肾上腺素与其受体结合后,促进偶联G蛋白构象改变。形成具有活性的GTP-Gsa, 而GTP.Gsa进一步激活腺苷酸环化酶(AC),激活的AC催化ATP形成第二信使cAMP(在细胞内浓度 可大10-5m0/几 总之,肾上腺素介导肝糖原和肌糖原的分解是通过以下信号转导通路进行的:首先,肾上腺素与B 肾上腺素受体结合,导致受体活性构象变化:激活后的受体进而与G蛋白结合,导致Gsα亚单位与G邱y 亚单位分离:同时Gsa与GDP的亲和力下降,与GTP的亲和力增加,形成Gsa-GTP复合物,G蛋白分子 开关打开;Gsa-CTP结合并激活AC,进而分解ATP,形成第二信使cAMP。cAMP激活蛋白激酶A (PKA),PKA进一步激活磷酸化酶激酶(phosphorylase kinase)的磷酸化;激活的磷酸化酶激酶进一步激 活糖原磷酸化酶,最终催化糖原降解为葡萄糖1-磷酸,再转化为葡萄糖-6-磷酸、葡萄糖(图20-8)。 在上述的时程中,虽然只有(100≈10)mo/几的肾上腺素被结合,但是能产生5mmol/L的葡错 糖。这说明反应过程中的激素的信号被逐级放大了约300万倍。也就是说,激素与受体结合后,可以在 几秒钟之内使磷酸化酶的活性达到最大值。 在不同的组织或细胞中,依赖cAMP的蛋白激酶A的底物是不相同的,cAMP通过活化或抑制不同 的酶系统,使细胞对不同的外界信号产生的生物学效应也是不同的。例如,肾上腺素通过©AMP和PKA 对糖原代谢的调控主要表现在肝脏和肌肉细胞。而在脂防细胞中,肾上腺素使PKA激活可以促讲磷脂 酶的磷酸化,磷酸化的磷脂酶进一步催化储存的甘油三酯水解,产生游离的脂肪酸和甘油分子,这些脂 肪酸将被释放到血液中被其他组织(如肾脏、心脏以及肌肉)作为能量来源摄取。 研究发现,不同的细胞中cAMP产生生物学效应时间是不同的。在肌肉细胞,1秒钟内可以启动糖 原降解为葡萄糖1-磷酸,而抑制糖原合成。而某些分泌细胞中,cAMP通过PKA产生生物学效应则需要 几个小时。已知PKA由两个调节亚基(R)和两个催化亚基(C)组成,每个R亚基有两个特异性的 AMP结合位点,当cAMP与PKA的调节亚基结合,改变调节亚基构象,使调节亚基和催化亚基解离,释 473