
生物大分子波 吴季 t1/2 decouple 1=x,2xy;3y,-X;4y,xXy;6-y,-xφef=X-X。每当增加一个值,1及接收机相 位反转.每个t1点采集二个fd,采集第二个fid时的相循环同第一个类似,但2-x,y:3y,x。 用第二步循环的相位: 21.S,sin()+21S,cos(sh -21,8,cos(h )-21,5.sin(h) ttr→-1,sin(24)-6Mo6in2I,S,cos(2g4) 2影,-L,9in(25)6e621,Scos(Q,4) 西ts→-EC.cos(2g4)+El,sin(Q,f) ,-8ed1.cos(Q4)+8l,sin(2,4) -a→+EMcI.cos(24)-£1,sin(Qg4) 两者相加得: (E:+Eueoun)l,sin(sh)+(8:+Eveon)I,cos(sh
PEP-HSQC 生物大分子波谱学原理 吴季辉
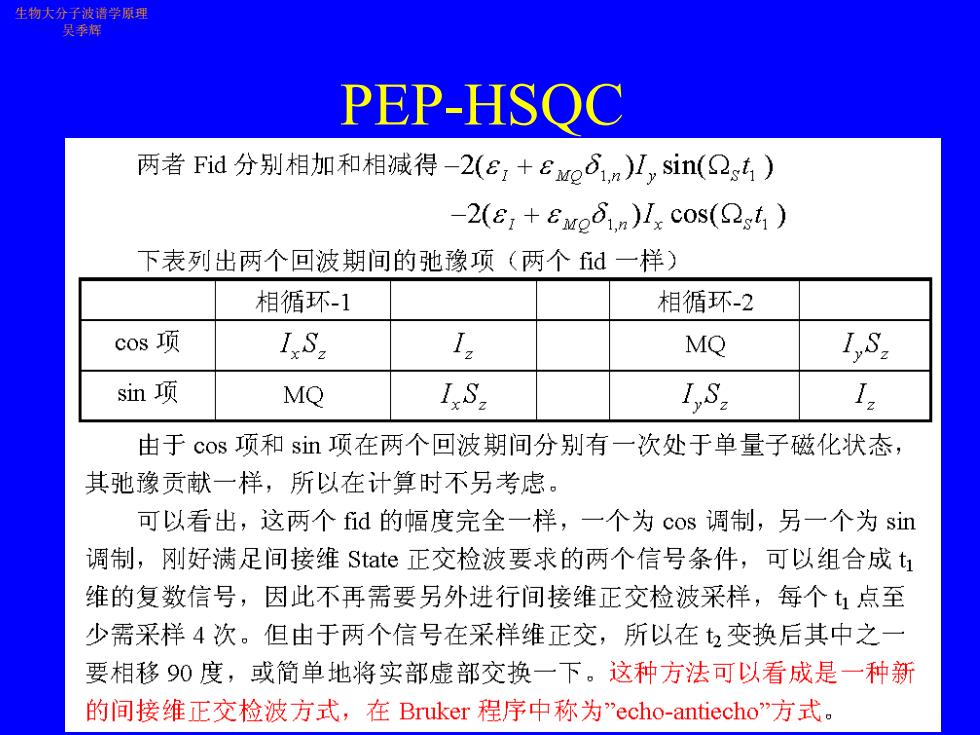
生物大分子波谱学原理 吴季辉 PEP-HSOC 两者Fid分别相加和相减得-2(e,+Eo6im)H,sin(2g4) 2(E,+Even)I,cos(sh 下表列出两个回波期间的弛豫项(两个fid一样) 相循环-1 相循环-2 cos项 1S2 I MQ I,S. sin项 MQ I.S. 1,S. I. 由于cos项和sn项在两个回波期间分别有一次处于单量子磁化状态, 其弛豫贡献一样,所以在计算时不另考虑。 可以看出,这两个fid的幅度完全一样,一个为cos调制,另一个为sin 调制,刚好满足间接维State正交检波要求的两个信号条件,可以组合成t1 维的复数信号,因此不再需要另外进行间接维正交检波采样,每个:点至 少需采样4次。但由于两个信号在采样维正交,所以在2变换后其中之一 要相移90度,或简单地将实部虚部交换一下。这种方法可以看成是一种新 的间接维正交检波方式,在Bruker程序中称为”echo-antiecho方式
PEP-HSQC 生物大分子波谱学原理 吴季辉

生物大分子波谐学原理 吴季辉 PEP-HSQC 3.PFG-PEP-HSQC 通常称为”gradient-enhanced HSQC”, 梯度场脉冲可以用于相干传递途径的选择, 但经常导致一半信号的损失,因为梯度场脉 冲选择信号依据信号的绝对相干阶。但是梯 度场脉冲可以同PEP-HSQC结合起来,既保 留正交的两个分量,又没有一半传递途径被 抑制的问题
PEP-HSQC 生物大分子波谱学原理 吴季辉 3. PFG-PEP-HSQC 通常称为”gradient-enhanced HSQC”, 梯度场脉冲可以用于相干传递途径的选择, 但经常导致一半信号的损失,因为梯度场脉 冲选择信号依据信号的绝对相干阶。但是梯 度场脉冲可以同PEP-HSQC结合起来,既保 留正交的两个分量,又没有一半传递途径被 抑制的问题

生物大分子波谱学原理 吴季辉 PEP-HSOC 2 Φ1 Φ1 t/2 t/2 decouple GZ 96 上图是PFG-PEP-HSQC的脉冲序列,其中2x= 1,2及接收 机相位均为x。每当1增加一个值,1及接收机相位反转。每个t1点采集二 个fid,采集第二个fid时2反转,同时s变号(或3)
PEP-HSQC 生物大分子波谱学原理 吴季辉

生物大分子波谐学原理 吴季辉 PEP-HSOC 图中a点之前序列同常规H$QC,有用信号是S的两个反相磁化 2IS,及2I2Sx。同样考虑InS体系,进一步的演化需考虑梯度场的 作用。 z方向梯度场使样品管不同高度z处的核受到的磁场强度变化, 因而进动频率变化:0,=⊙。-zG,第一项为正常的化学位移, 在回波序列中可以不考虑,后一项为梯度的贡献,产生的相移为 中2=zG2t,此处t是梯度的作用时间。从Hamiltonian角度讲,梯 度场的作用相当于绕z轴的旋转,角度为中
PEP-HSQC 生物大分子波谱学原理 吴季辉