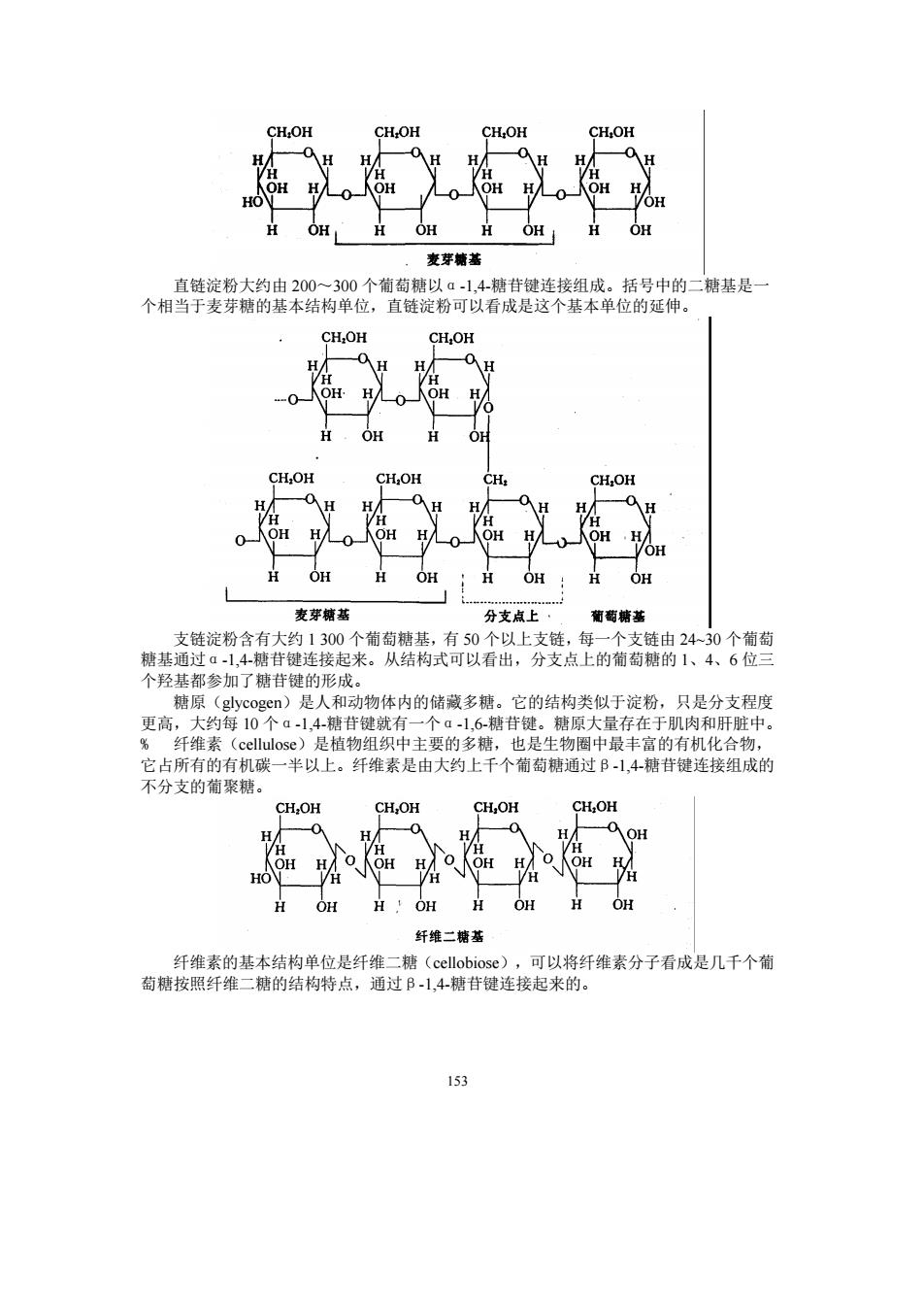
CH:OR ,数芽塘著 00个葡萄 二糖基是 CHOH CH. CH,OH H OH H OH H OH H OH 麦芽糖基 分支直上, 个羟基都参加了糖苷健的形成。 糖原(glycogen)是人和动物体内的储藏多糖。它的结构类似于淀粉,只是分支程度 更高,大约每10个a1,4糖苷键就有一个a1,6糖苷键。糖原大量存在于肌肉和肝脏中。 %纤维素(cellulos心)是植物组织中主要的多糖,也是生物圈中最丰富的有机化合物, 它占所有的有机碳一半以上。纤维素是由大约上千个葡萄糖通过B-1,4糖苷键连接组成的 不分支的萄聚糖。 CH:OH CH,OH H H人 OH H H OH H OH H OH 纤维二精基 (cellobiose 可以将纤维素分子看成是几千个葡 萄糖按照纤维 153
153 直链淀粉大约由 200~300 个葡萄糖以α-1,4-糖苷键连接组成。括号中的二糖基是一 个相当于麦芽糖的基本结构单位,直链淀粉可以看成是这个基本单位的延伸。 支链淀粉含有大约 1 300 个葡萄糖基,有 50 个以上支链,每一个支链由 24~30 个葡萄 糖基通过α-1,4-糖苷键连接起来。从结构式可以看出,分支点上的葡萄糖的 1、4、6 位三 个羟基都参加了糖苷键的形成。 糖原(glycogen)是人和动物体内的储藏多糖。它的结构类似于淀粉,只是分支程度 更高,大约每 10 个α-1,4-糖苷键就有一个α-1,6-糖苷键。糖原大量存在于肌肉和肝脏中。 % 纤维素(cellulose)是植物组织中主要的多糖,也是生物圈中最丰富的有机化合物, 它占所有的有机碳一半以上。纤维素是由大约上千个葡萄糖通过β-1,4-糖苷键连接组成的 不分支的葡聚糖。 纤维素的基本结构单位是纤维二糖(cellobiose),可以将纤维素分子看成是几千个葡 萄糖按照纤维二糖的结构特点,通过β-1,4-糖苷键连接起来的

第二节双糖和多糖的酶促降解 生物体中的双糖和多糖都是在相应醇的催化下被水解的 一、双糖的酶促降解% )芯糖的水解 蔗糖的水解由蔗糖酶催化,此酶也称转化酶(invertase),在植物体内广泛存在。蔗 糖水解后产生1分子葡萄糖和1分子果糖。 CHOH CHOH ,0 HoCH,O、H H OI CH,OH OH OH H H OH 。一D一龙喇葡药糖 (二)麦芽糖的水解 麦芽糖酶催化1分子麦芽糖水解产生2分子a-D-葡萄糖。另外,植物中还存在α-葡 萄糖苷醇,此酶也可催化麦芽糖的水解,在含淀粉种子萌发时最丰富。 CHOH CHOH H HO OH H OH H OH 芽糖 &一D一葡萄辅 (三)乳糖的水解 糖的水解由乳糖南催化,生成1分子半乳糖和1分子葡萄糖 CH,OH HO 乳塘 一吡嗜葡萄 154
154 第二节 双糖和多糖的酶促降解 生物体中的双糖和多糖都是在相应酶的催化下被水解的。 一、双糖的酶促降解% 生物体中的双糖在相应酶的催化下被降解为单糖,然后进一步被氧化分解,或转化为 其它化合物。例如,人和高等动物的肠粘膜细胞中有蔗糖酶、乳糖酶和麦芽糖酶,可以将 相应的双糖降解。 (一)蔗糖的水解 蔗糖的水解由蔗糖酶催化,此酶也称转化酶(invertase),在植物体内广泛存在。蔗 糖水解后产生 1 分子葡萄糖和 1 分子果糖。 (二)麦芽糖的水解 麦芽糖酶催化 1 分子麦芽糖水解产生 2 分子α-D-葡萄糖。另外,植物中还存在α-葡 萄糖苷酶,此酶也可催化麦芽糖的水解,在含淀粉种子萌发时最丰富。 (三)乳糖的水解 乳糖的水解由乳糖酶催化,生成 1 分子半乳糖和 1 分子葡萄糖

二、淀粉的酶促降解 (一)淀粉的水解 能够催化淀粉a-1,4糖苷键以及a-1,6-糖苷键水解的酶叫淀粉酶(amylase),主要包 括ā-淀粉酶、B-淀粉酶以及R酶。 1.a-淀粉碑 a-淀粉醇又称a-l,4葡聚糖水解酵。这是一种内切淀粉酶(endoamylase),可以水 解直链淀粉或糖原分子内部的任意α-1,4糖苷键,但对距淀粉链非还原性末端第五个以后 水解产物为葡萄糖和麦芽糖 、麦芽三糖以 09 6- 的湿物 2.B淀粉 B-淀粉酶又称a-1,4-葡聚糖基麦芽糖基水解酶。这是一种外切淀粉酶(x0 从淀粉分子外围的非还原性末端开始,每间隔 个进行解生型物为 如果底物是直链淀粉,水解产物几乎都是麦芽糖:如果底物是支链淀粉,水解产物为麦芽 糖和多分支糊精(B极限糊精)。 ā-淀粉酶是需要与Ca结合而表现活性的金属酶,因此整合剂EDTA等能抑制此酶 B淀粉酶是含巯基的酶,氧化疏基的试剂能抑制此酶。ā-淀粉酶耐热不耐酸,在pH3.3 中的和B并不是指其作用的 或B苷钟 不同的正 际上,这两种酶都只 粉的a1,4 苷健,水解的终产物以麦芽糖为主(图5-3) 图5一3。一淀粉酶和一淀粉离对支链淀粉的分解作用 3.R-酶 R-酶又称为脱支酶(debranching enzyme),它可作用于淀粉的a-l,6-糖苷键,但它不 能水解支链淀粉内部的分支,只能水解支链淀粉的外围分支。所以,支链淀粉的完全降解 155
155 二、淀粉的酶促降解 (一)淀粉的水解 能够催化淀粉α-1,4-糖苷键以及α-1,6-糖苷键水解的酶叫淀粉酶(amylase),主要包 括α-淀粉酶、β-淀粉酶以及 R-酶。 1. α-淀粉酶 α-淀粉酶又称α-1,4-葡聚糖水解酶。这是一种内切淀粉酶(endoamylase),可以水 解直链淀粉或糖原分子内部的任意α-1,4-糖苷键,但对距淀粉链非还原性末端第五个以后 的糖苷键的作用受到抑制。当底物是直链淀粉,水解产物为葡萄糖和麦芽糖、麦芽三糖以 及低聚糖的混合物;当底物是支链淀粉,则直链部分的α-1,4-糖苷键被水解,而α-1,6-糖 苷键不被水解,水解产物为葡萄糖和麦芽糖、麦芽三糖等寡聚糖类以及含有α-1,6-糖苷键 的短的分支部分极限糊精(α-极限糊精)的混合物。 2. β-淀粉酶 β-淀粉酶又称α-1,4-葡聚糖基-麦芽糖基水解酶。这是一种外切淀粉酶(exoamylase), 从淀粉分子外围的非还原性末端开始,每间隔一个糖苷键进行水解,生成产物为麦芽糖。 如果底物是直链淀粉,水解产物几乎都是麦芽糖;如果底物是支链淀粉,水解产物为麦芽 糖和多分支糊精(β-极限糊精)。 α-淀粉酶是需要与 Ca +结合而表现活性的金属酶,因此螯合剂 EDTA 等能抑制此酶。 β-淀粉酶是含巯基的酶,氧化巯基的试剂能抑制此酶。α-淀粉酶耐热不耐酸,在 pH3.3 时酶被破坏,而在 70℃下,保持 15min 该酶仍保持活性。β-淀粉酶则耐酸不耐热,在 pH3.3 时酶可保持活性,但在 70℃下 15min 酶被破坏。 要需说明的是:α-淀粉酶和β-淀粉酶中的α和β并不是指其作用的α-或β-糖苷键, 而只是表明对淀粉水解作用不同的两种酶,实际上,这两种酶都只作用于淀粉的α-1,4-糖 苷键,水解的终产物以麦芽糖为主(图 5-3)。 3. R-酶 R-酶又称为脱支酶(debranching enzyme),它可作用于淀粉的α-1,6-糖苷键,但它不 能水解支链淀粉内部的分支,只能水解支链淀粉的外围分支。所以,支链淀粉的完全降解

需要有ā-淀粉酶、B-淀粉酶和R-酶的共同作用。 群 ,也可以被磷酸解(phosphorolysis)。 降 生成的1一磷酸葡萄糖不 萄糖变位酶的催化下转化为6碳酸葡萄糖,最后转化为葡萄糖,6-磷酸葡萄糖也可直接经 链接解被氨化。 由于磷酸化酶只能作用于α-14糖苷键,所以不能完全降解支链淀粉,支 链淀粉的完全降解还需有其它酶的配合。 2a-16-糖苷键的降解 支链淀粉经过磷酸解完全降解需三种酶的共同作用。这三种酶是磷酸化酶、转移酶和 a-1,6-糖苷酶。首先,磷酸化酶(phosphorylase)从非还原性末端依次降解并释放出1分 子1磷酸葡萄糖,直到在分支点以前还剩4个葡萄糖残基为止。 然后转移酶(transferase 的4个萄糖残基种的 个葡萄糖残基转移到另 并形 糖苷 ,原来的分文结构变成了直结构瑞酸化鹅可维续性化酸解,生成 糖原的降解也是通过磷酸解,由磷酸化酶和转移酶以及α-1,6-糖苷酶共同作用将糖原 完全降解。整个过程见图5-4。$ 0-0-0-0-0-e台 0-00-06 化 (拜救8个G-1-P) 只9-。g衔 转移 999e9R9999 脱支酶 9999998896891 G-1P(多个) 图54支链淀粉(或糖原)彻底磷酸解的步骤 156
156 需要有α-淀粉酶、β-淀粉酶和 R-酶的共同作用。 (二)淀粉的磷酸解 淀粉除了可以被水解外,也可以被磷酸解(phosphorolysis)。 1. α-1,4-糖苷键的降解 淀粉磷酸化酶可作用于淀粉的α-1,4-糖苷键,从非还原端依次进行磷酸解,每次释放 1 分子 1-磷酸葡萄糖。生成的 1-磷酸葡萄糖不能扩散到细胞外, 并且可进一步在磷酸葡 萄糖变位酶的催化下转化为 6-磷酸葡萄糖,最后转化为葡萄糖,6 -磷酸葡萄糖也可直接经 糖酵解被氧化。由于磷酸化酶只能作用于α-1,4-糖苷键,所以不能完全降解支链淀粉,支 链淀粉的完全降解还需有其它酶的配合。 2. α-1,6-糖苷键的降解 支链淀粉经过磷酸解完全降解需三种酶的共同作用。这三种酶是磷酸化酶、转移酶和 α-1,6-糖苷酶。首先,磷酸化酶(phosphorylase)从非还原性末端依次降解并释放出 1 分 子 1-磷酸葡萄糖,直到在分支点以前还剩 4 个葡萄糖残基为止。然后转移酶(transferase) 将一个分支上剩下的 4 个葡萄糖残基中的 3 个葡萄糖残基转移到另一个分支上,并形成一 个新的α-1,4-糖苷键。最后,α-1,6-糖苷酶(α-1,6-glucosidase)降解暴露在外的α-1,6- 糖苷键。这样,原来的分支结构就变成了直链结构,磷酸化酶可继续催化其磷酸解,生成 1-磷酸葡萄糖。 糖原的降解也是通过磷酸解,由磷酸化酶和转移酶以及α-1,6-糖苷酶共同作用将糖原 完全降解。整个过程见图 5-4。$ 图 5-4 支链淀粉(或糖原)彻底磷酸解的步骤

三、纤维素的酶促降解 纤维素是由B山,4葡萄糖苷键组成的多糖,虽然也以葡萄糖为基本组成单位,但其性 质与淀粉有很大差异,纤维素是一种结枸多糖而不起营养作用 纤维素的降解是在纤维素酶(cellulase)的催化下进行的。有些微生物(包括真菌、 放线菌、细菌)及反刍动物的消化系统瘤胃中的某些细菌能产生纤维素酶,所以能降解与 消化纤维素。而哺乳动物没有纤维素酶,所以不能消化植物纤维。 纤维素酶是参与水解纤维素的一类酶的总称,采用各种层析和电泳技术等可将纤维素 酶分成不同的 组分,主要包括C酶、C,酶和B-葡萄糖苷酶三种类型。 维素品 酶系中的 它在天然纤维 的降解过程中起主导作用。C酶破 构,使其 可被 C、酶也称B-1,4葡聚糖酶,是水解酶,能水解溶解的纤维素衍生物或者膨胀和部分降 解的纤维素,但不能作用于结晶的纤维素 B-1,4葡聚糖酶有二种类型:外切B-1,4葡聚糖酶和内切B1,4葡聚糖酶 外切B1,4葡聚糖酶能从纤维素链的非还原性末端一个一个地依次切下葡萄糖单位, 物是ā-简萄糖。专一性比较强。 它对纤维寡糖的亲合力强,能迅速水解内切酶作用产 生的纤维真糖。 系统命名为B-1,4葡聚糖葡萄糖水解酶(EC32.1.- 内切B1,4葡聚糖酶以随机形式水解 14葡聚糖, 是纤维糊精、纤 系统命名为书1.4 EC32.1.4)。 (EC32.121)也称纤维二糖酶,能水解纤维二糖和短健的纤维寡糖生 成葡萄糖。对纤维二糖和纤维三糖的水解很快,随着葡萄糖聚合度的增加水解速度下降, 它水解纤维二糖生成2分子葡萄糖。 在这上述几种酶的共同作用下,纤维素可被水解为葡萄糖: 晶状天然纤维素 9 无定型游离纤维素C B-葡萄糖苷 一纤维一塘 葡萄糖 第三节糖酵解 是生物体取得能量的主要方 。糖的分解就是指糖的氧 生物体中糖 的 是葡 的进行, 是所有生物体进行葡萄糖分解代谢所必须经过的共同阶段】 糖酵解作用的阐明,主要依赖动物肌肉和酵母的实验结果,其全过程于1940年就已 弄清楚,在这项研究中,有三位德国生物化学家:Gustav Embden、Otto Meyerhof、Jacob Parnas的贡献最大,因此,糖酵解过程又称为Embden-Meyerhof-Pamas途径,简称EMP 途径。釉酵解作用进行的场所是细胞质
157 三、纤维素的酶促降解 纤维素是由β-1,4-葡萄糖苷键组成的多糖,虽然也以葡萄糖为基本组成单位,但其性 质与淀粉有很大差异,纤维素是一种结构多糖而不起营养作用。 纤维素的降解是在纤维素酶(cellulase)的催化下进行的。有些微生物(包括真菌、 放线菌、细菌)及反刍动物的消化系统瘤胃中的某些细菌能产生纤维素酶,所以能降解与 消化纤维素。而哺乳动物没有纤维素酶,所以不能消化植物纤维。 纤维素酶是参与水解纤维素的一类酶的总称,采用各种层析和电泳技术等可将纤维素 酶分成不同的组分,主要包括 C1 酶、Cx酶和β-葡萄糖苷酶三种类型。 (一、)C1酶 C1 酶是纤维素酶系中的重要组分,它在天然纤维素的降解过程中起主导作用。C1酶破 坏天然纤维素晶状结构,使其变成可被 Cx酶作用的形式。 (二)Cx酶 Cx酶也称β-1,4-葡聚糖酶,是水解酶,能水解溶解的纤维素衍生物或者膨胀和部分降 解的纤维素,但不能作用于结晶的纤维素。 β-1,4-葡聚糖酶有二种类型:外切β-1,4-葡聚糖酶和内切β-1,4-葡聚糖酶 外切β-1,4-葡聚糖酶能从纤维素链的非还原性末端一个一个地依次切下葡萄糖单位, 产物是α-葡萄糖。专一性比较强。 它对纤维寡糖的亲合力强,能迅速水解内切酶作用产 生的纤维寡糖。系统命名为β-1,4-葡聚糖葡萄糖水解酶(EC 3.2.1.-) 内切β-1,4-葡聚糖酶以随机形式水解β-1,4-葡聚糖,它作用于较长的纤维素链,对末 端键的敏感性比间键小,主要产物是纤维糊精、纤维二糖和纤维三搪。系统命名为β-1,4- 葡聚糖葡聚糖水解酶(EC 3.2.1.4)。 (三)β-葡萄糖苷酶 β-葡萄糖苷酶(EC 3.2.1.21)也称纤维二糖酶,能水解纤维二糖和短键的纤维寡糖生 成葡萄糖。对纤维二糖和纤维三糖的水解很快,随着葡萄糖聚合度的增加水解速度下降。 它水解纤维二糖生成 2 分子葡萄糖。 在这上述几种酶的共同作用下,纤维素可被水解为葡萄糖: C1 酶 Cx酶 β-葡萄糖苷酶 晶状天然纤维素 无定型游离纤维素 纤维二糖 葡萄糖 第三节 糖酵解 糖的分解代谢是生物体取得能量的主要方式。糖的分解就是指糖的氧化。生物体中糖 的氧化分解主要有三条途径:糖的无氧氧化、糖的有氧氧化和磷酸戊糖途径。其中糖的无 氧氧化又称糖酵解(glycolysis)。糖酵解是葡萄糖经 1,6-二磷酸果糖和 3-磷酸甘油酸转变 为丙酮酸,同时产生 ATP 的一系列反应。这一过程无论在有氧或厌氧的条件下均可进行, 是所有生物体进行葡萄糖分解代谢所必须经过的共同阶段。 糖酵解作用的阐明,主要依赖动物肌肉和酵母的实验结果,其全过程于 1940 年就已 弄清楚,在这项研究中,有三位德国生物化学家:Gustav Embden、Otto Meyerhof、Jacob Parnas 的贡献最大,因此,糖酵解过程又称为 Embden-Meyerhof-Parnas 途径,简称 EMP 途径。糖酵解作用进行的场所是细胞质