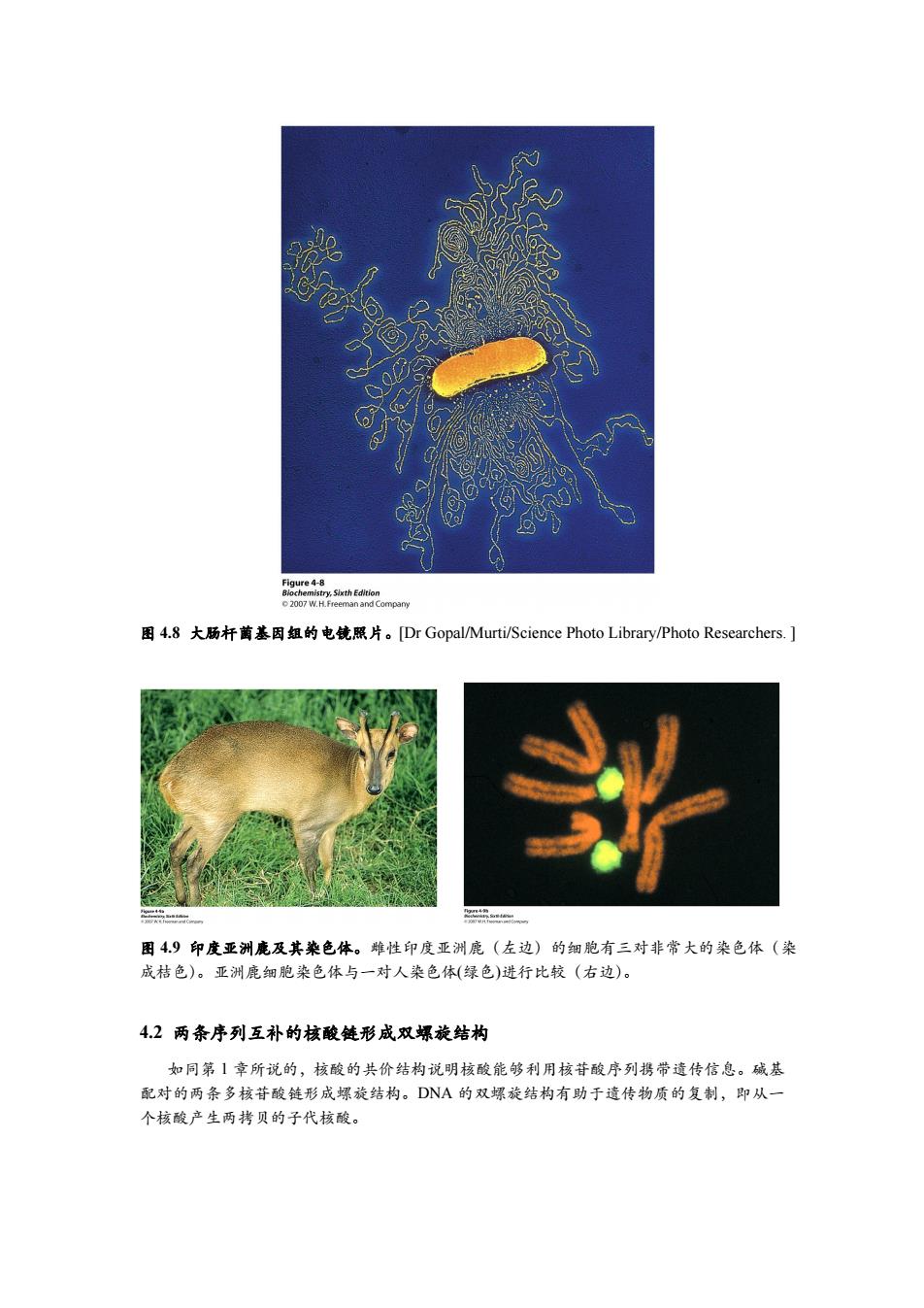
0Q② Figure 4-8 图4.8大肠杆菌基因组的电镜照片。[Dr Gopal/Murti/Science Photo Library/Photo Researchers.] 图49印度亚洲鹿及其染色体。雌性印度亚洲鹿(左边)的细胞有三对非常大的染色体(染 成桔色)。亚洲鹿细胞染色体与一对人染色体(绿色)进行比较(右边)。 4.2两条序列互补的核酸链形成双螺旋结构 如同第1章所说的,核酸的共价结构说明核酸能够利用核苷酸序列携带遗传信息。碱基 配对的两条多核苷酸链形成螺旋结构。DNA的双螺旋结构有助于遗传物质的复制,即从一 个核酸产生两拷贝的子代核酸
图 4.8 大肠杆菌基因组的电镜照片。[Dr Gopal/Murti/Science Photo Library/Photo Researchers. ] 图 4.9 印度亚洲鹿及其染色体。雌性印度亚洲鹿(左边)的细胞有三对非常大的染色体(染 成桔色)。亚洲鹿细胞染色体与一对人染色体(绿色)进行比较(右边)。 4.2 两条序列互补的核酸链形成双螺旋结构 如同第 1 章所说的,核酸的共价结构说明核酸能够利用核苷酸序列携带遗传信息。碱基 配对的两条多核苷酸链形成螺旋结构。DNA 的双螺旋结构有助于遗传物质的复制,即从一 个核酸产生两拷贝的子代核酸
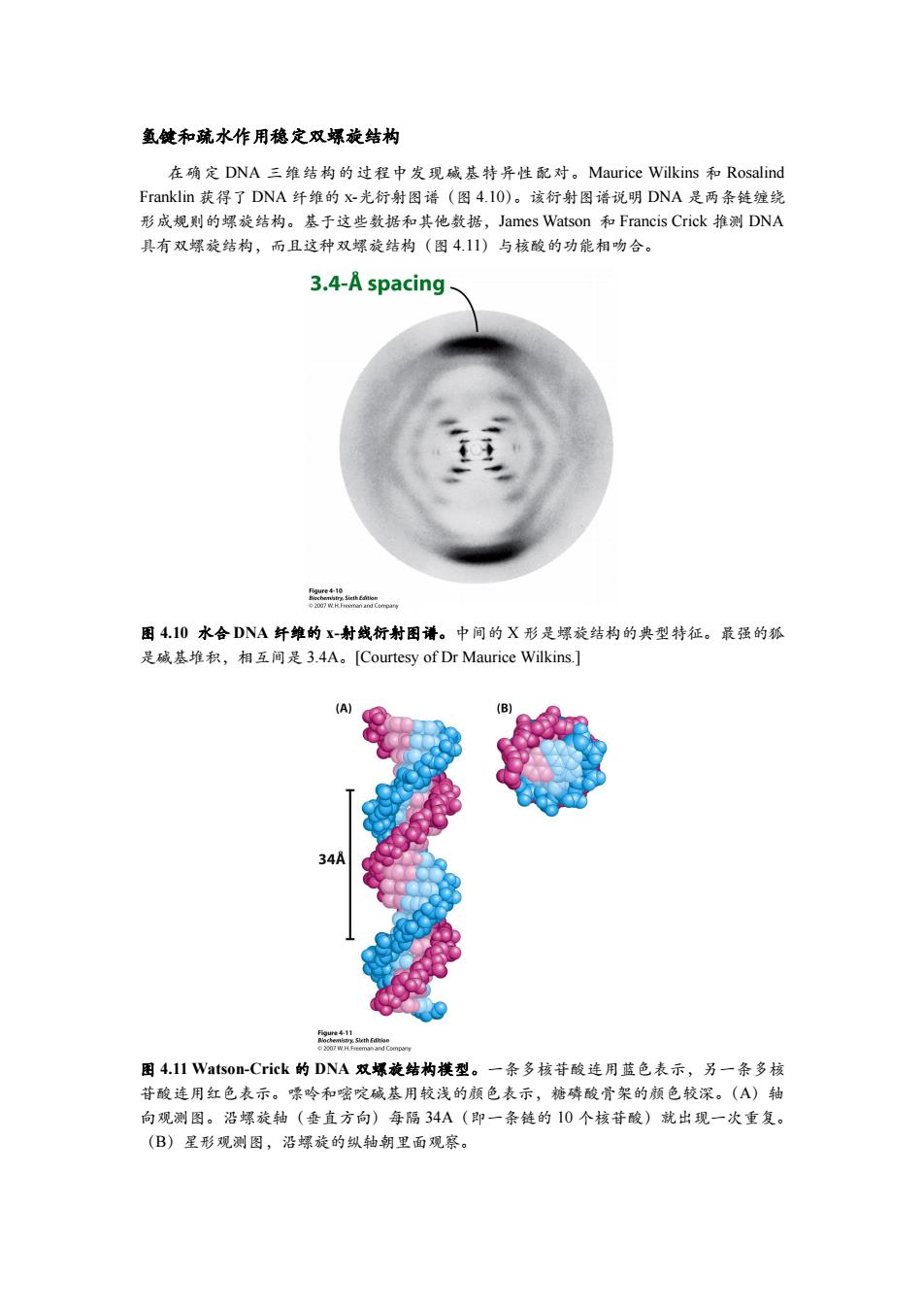
氢健和疏水作用稳定双螺旋结构 在确定DNA三维结构的过程中发现碱基特异性配对。Maurice Wilkins和Rosalind Franklin获得了DNA纤维的X-光衍射图谱(图4.l0)。该衍射图谱说明DNA是两条链缠绕 形成规则的螺旋结构。基于这些数据和其他数据,James Watson和Francis Crick推测DNA 具有双螺旋结构,而且这种双螺旋结构(图4.11)与核酸的功能相吻合。 3.4-A spacing en 图4.10水合DNA纤维的X-射线衍射图谱。中间的X形是螺旋结构的典型特征。最强的狐 是碱基堆积,相互间是3.4A。[Courtesy of Dr Maurice Wilkins.] (A) (B) 34A 图4.l1 Vatson-Crick的DNA双螺旋结构樸型。一条多核苷酸连用蓝色表示,另一条多核 苷酸连用红色表示。嘌呤和嘧啶碱基用较浅的颜色表示,糖磷酸骨架的颜色较深。(A)轴 向观测图。沿螺旋轴(垂直方向)每隔34A(即一条链的10个核苷酸)就出现一次重复。 (B)星形观测图,沿螺旋的纵轴朝里面观察
氢键和疏水作用稳定双螺旋结构 在确定 DNA 三维结构的过程中发现碱基特异性配对。Maurice Wilkins 和 Rosalind Franklin 获得了 DNA 纤维的 x-光衍射图谱(图 4.10)。该衍射图谱说明 DNA 是两条链缠绕 形成规则的螺旋结构。基于这些数据和其他数据,James Watson 和 Francis Crick 推测 DNA 具有双螺旋结构,而且这种双螺旋结构(图 4.11)与核酸的功能相吻合。 图 4.10 水合 DNA 纤维的 x-射线衍射图谱。中间的 X 形是螺旋结构的典型特征。最强的狐 是碱基堆积,相互间是 3.4A。[Courtesy of Dr Maurice Wilkins.] 图 4.11 Watson-Crick 的 DNA 双螺旋结构模型。一条多核苷酸连用蓝色表示,另一条多核 苷酸连用红色表示。嘌呤和嘧啶碱基用较浅的颜色表示,糖磷酸骨架的颜色较深。(A)轴 向观测图。沿螺旋轴(垂直方向)每隔 34A(即一条链的 10 个核苷酸)就出现一次重复。 (B)星形观测图,沿螺旋的纵轴朝里面观察

James Watson和Francis Crick从x-光衍射图谱推测的DNA双螺旋结构具有下列特征: 1.两条核酸链方向相反,围绕同一轴缠绕形成螺旋结构。 2.糖-磷酸骨架在双螺旋外侧,碱基处于双螺旋内部。 3.碱基几乎与双螺旋轴垂直,相邻碱基间的距离是3.4A。这种间距在X光衍射图谱(图 4.10)中可以看出。一个螺旋结构重复是34A,10个碱基。因此每个碱基旋转36°。 4.螺旋的直径是20A。 如果嘌吟和嘧啶的大小和形状不同,这种规则结构如何适应任一序列?为了回答这个问 题,James Watson和Francis Crick发现鸟嘌呤与胞嘧啶配对和腺嘌吟与胸腺嘧啶配对的形 状几乎相同(图412)。碱基之间特异的氢键将碱基结合在一起形成碱基对。尽管单一氢键 的键能只有4~21kJ/mol(或1~5kcal/mol),但是DNA分子有大量氢键产生了可观的结合 力。这种碱基配对模式能够解释Erwin Chargaft在I950年观察的现象,即所有DNA分子中 腺嘌吟与胸腺嘧啶的比例和鸟嘌呤与胞嘧啶的比例几乎相同,而腺嘌吟与鸟嘌吟的比值在不 同物种中差异很大(表4.1)。 -A Guanine Cytosine Adenine Thymine n 图4.l2 Watson和Crick提出的配对碱基结构。 TABLE 4.I Base compositions experimentally determined for a variety of organisms Species A:T G:C A:G Human being 1.00 1.00 1.56 Salmon 1.02 1.02 1.43 Wheat 1.00 0.97 1.22 Yeast 1.03 1.02 1.67 Escherichia coli 1.09 0.99 1.05 Serratia marcescens 0.95 0.86 0.70 Table 4-1 Biochemistry,Sixth Edition 2007 W.H.Freeman and Company
James Watson 和 Francis Crick 从 x-光衍射图谱推测的 DNA 双螺旋结构具有下列特征: 1. 两条核酸链方向相反,围绕同一轴缠绕形成螺旋结构。 2. 糖-磷酸骨架在双螺旋外侧,碱基处于双螺旋内部。 3. 碱基几乎与双螺旋轴垂直,相邻碱基间的距离是 3.4A。这种间距在 x-光衍射图谱(图 4.10)中可以看出。一个螺旋结构重复是 34A,10 个碱基。因此每个碱基旋转 36 o。 4. 螺旋的直径是 20A。 如果嘌呤和嘧啶的大小和形状不同,这种规则结构如何适应任一序列?为了回答这个问 题,James Watson 和 Francis Crick 发现鸟嘌呤与胞嘧啶配对和腺嘌呤与胸腺嘧啶配对的形 状几乎相同(图 4.12)。碱基之间特异的氢键将碱基结合在一起形成碱基对。尽管单一氢键 的键能只有 4 ~ 21 kJ/mol(或 1 ~ 5 kcal/mol),但是 DNA 分子有大量氢键产生了可观的结合 力。这种碱基配对模式能够解释 Erwin Chargaff 在 1950 年观察的现象,即所有 DNA 分子中 腺嘌呤与胸腺嘧啶的比例和鸟嘌呤与胞嘧啶的比例几乎相同,而腺嘌呤与鸟嘌呤的比值在不 同物种中差异很大(表 4.1)。 图 4.12 Watson 和 Crick 提出的配对碱基结构

在双螺旋内部,一层碱基对堆积在另一层碱基对上。碱基堆积对双螺旋稳定性的贡献表 现在两个方面。碱基堆积构成的疏水效应稳定了DNA双螺旋结构。疏水碱基聚集于双螺旋 内部(避开DNA的水环境),而双螺旋表面更亲水。双螺旋结构的这种排列与蛋白质折叠 很相似(蛋白质折叠将疏水残基包裹在分子内部、亲水残基暴露于分子外部)(2.4节)。疏 水效应使碱基推积于另一个碱基之上。堆积碱基之间的吸引力是范德华力。单个范德华力很 弱,通常在2~4kJ/mol(0.5~1kca/mol)。然而,双螺旋核酸分子中大量原子处于范德华 接触状态,因此范德华的净效应,即这些原子相互作用力的总和非常显著。此外DNA分子 骨架的五元环糖基具有刚性结构,有利于碱基堆积。 图4.13DNA纤维的戴面图。在双螺旋内部一个碱基对几乎正好堆积在另一碱基对的顶部。 双螺旋结构有利于遗传信息的精确传递 DNA的双螺旋结构和碱基之间形成碱基对提示遗传物质是如何复制的。双螺旋中一条 链核苷酸序列能精确决定另一条互补链的核苷酸序列。一条链的鸟嘌呤总是与另一条链的胞 嘧啶配对,其余碱基配对均遵循这些法测。因此双螺旋DNA链分开后形成的两条链都可充 当模板指导新的双螺旋DNA的构建。结果,子代DNA分子有一条链是来自亲本,另一条 链是新合成的。这就是半保留复制。 1958年Mathew Meselson和Franklin Stahl的试验验证了这个假说。他们用15N标记亲 本DNA,因此DNA链比普通DNA链重。将大肠杆菌置于以15N标记的NH4CI作为唯一氦 源的培养基中生长多代,其DNA就完全为15N标记。然后将细菌转移到只含有14N作为氨 源的培养基中生长,看看15N和1N在DNA中的分布。 密度梯度平衡离心分析15N和1N在DNA中的分布。少量DNA分子溶于密度接近 DNA(I.7gmL)的CsCI溶液中。离心直至平衡状态。此时沉降和扩散这两个相反作用达到平 衡,在离心管中形成1.66~1.76gmL的密度梯度。此时,DNA分子被迫进入密度与自身密 度相等的介质中。离心导致基因组DNA形成很窄的条带,可以用紫外吸收观测。由于1N 和1N在DNA的密度差异大约是1%,密度梯度平衡离心能形成两条明显的条带(图4.14)
在双螺旋内部,一层碱基对堆积在另一层碱基对上。碱基堆积对双螺旋稳定性的贡献表 现在两个方面。碱基堆积构成的疏水效应稳定了 DNA 双螺旋结构。疏水碱基聚集于双螺旋 内部(避开 DNA 的水环境),而双螺旋表面更亲水。双螺旋结构的这种排列与蛋白质折叠 很相似(蛋白质折叠将疏水残基包裹在分子内部、亲水残基暴露于分子外部)(2.4 节)。疏 水效应使碱基堆积于另一个碱基之上。堆积碱基之间的吸引力是范德华力。单个范德华力很 弱,通常在 2 ~ 4 kJ/mol (0.5 ~ 1 kcal/mol)。然而,双螺旋核酸分子中大量原子处于范德华 接触状态,因此范德华的净效应,即这些原子相互作用力的总和非常显著。此外 DNA 分子 骨架的五元环糖基具有刚性结构,有利于碱基堆积。 图 4.13 DNA 纤维的截面图。在双螺旋内部一个碱基对几乎正好堆积在另一碱基对的顶部。 双螺旋结构有利于遗传信息的精确传递 DNA 的双螺旋结构和碱基之间形成碱基对提示遗传物质是如何复制的。双螺旋中一条 链核苷酸序列能精确决定另一条互补链的核苷酸序列。一条链的鸟嘌呤总是与另一条链的胞 嘧啶配对,其余碱基配对均遵循这些法则。因此双螺旋 DNA 链分开后形成的两条链都可充 当模板指导新的双螺旋 DNA 的构建。结果,子代 DNA 分子有一条链是来自亲本,另一条 链是新合成的。这就是半保留复制。 1958 年 Mathew Meselson 和 Franklin Stahl 的试验验证了这个假说。他们用 15N 标记亲 本 DNA,因此 DNA 链比普通 DNA 链重。将大肠杆菌置于以 15N 标记的 NH4Cl 作为唯一氮 源的培养基中生长多代,其 DNA 就完全为 15N 标记。然后将细菌转移到只含有 14N 作为氮 源的培养基中生长,看看 15N 和 14N 在 DNA 中的分布。 密度梯度平衡离心分析 15N 和 14N 在 DNA 中的分布。少量 DNA 分子溶于密度接近 DNA(1.7g/mL)的 CsCl 溶液中。离心直至平衡状态。此时沉降和扩散这两个相反作用达到平 衡,在离心管中形成 1.66 ~ 1.76g/mL 的密度梯度。此时, DNA 分子被迫进入密度与自身密 度相等的介质中。离心导致基因组 DNA 形成很窄的条带,可以用紫外吸收观测。由于 15N 和 14N 在 DNA 的密度差异大约是 1%,密度梯度平衡离心能形成两条明显的条带(图 4.14)
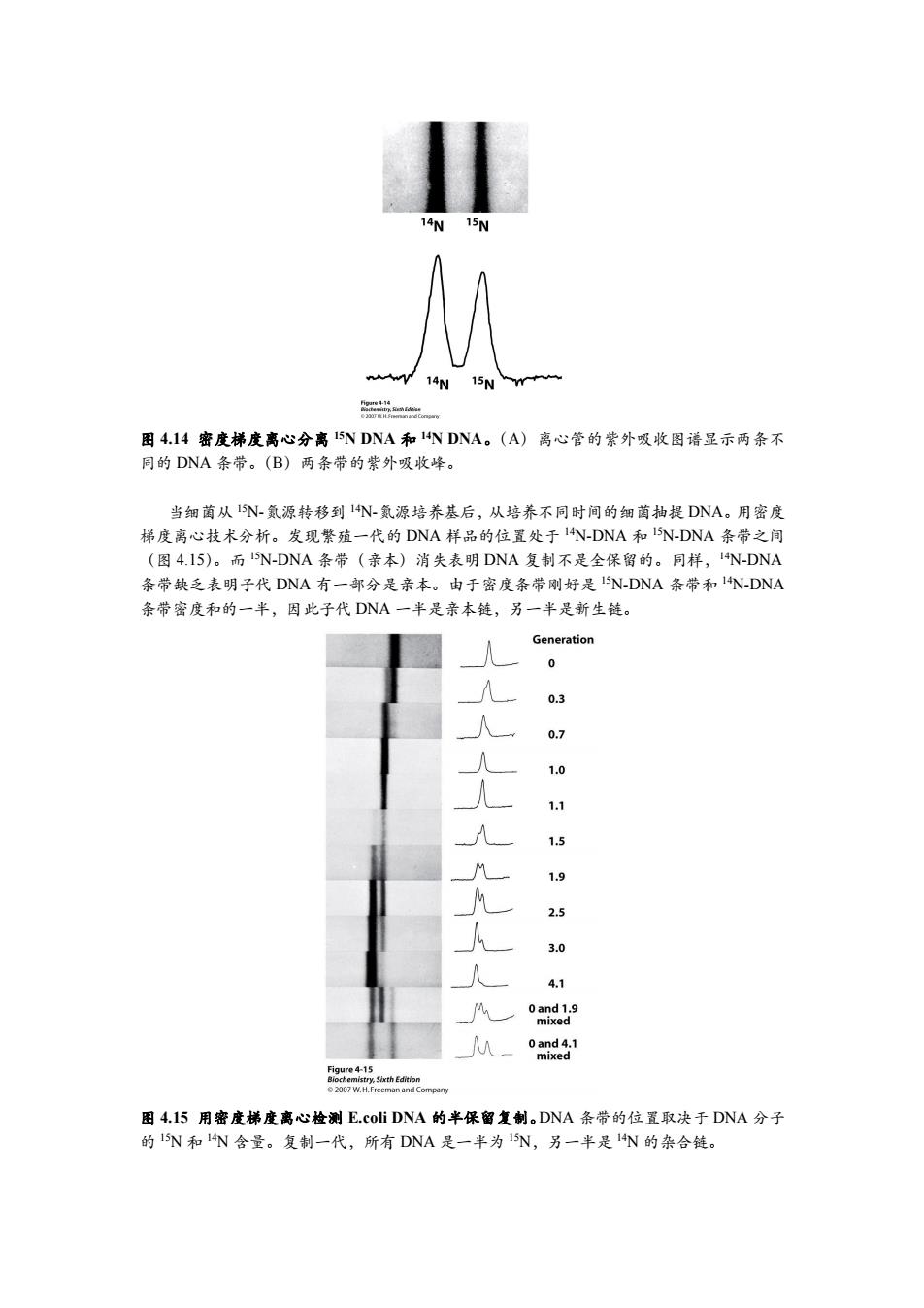
14W15N 14N 15Nm 图4.14密度梯度离心分离15NDNA和1 N DNA。(A)离心管的紫外吸收图谱显示两条不 同的DNA条带。(B)两条带的紫外吸收峰。 当细菌从15N-氦源转移到1N-氨源培养基后,从培养不同时间的细菌抽提DNA。用密度 梯度离心技术分析。发现繁殖一代的DNA样品的位置处于14N-DNA和I5N-DNA条带之间 (图4.I5)。而15N-DNA条带(亲本)消失表明DNA复制不是全保留的。同样,14N-DNA 条带缺乏表明子代DNA有一部分是亲本。由于密度条带刚好是I5N-DNA条带和1N-DNA 条带密度和的一半,因此子代DNA一半是亲本链,另一半是新生链。 Generation 0 0.7 1.0 1.1 1.5 九九人人 1.9 2.5 3.0 4.1 0and1.9 mixed 0 and 4.1 mixed Figure 4-15 Biochemistry.Sixth Edition 2007 W.H.Freeman and Company 图4.15用密度梯度离心检测E.coli DNA的半保留复制。DNA条带的位置取决于DNA分子 的15N和1N含量。复制一代,所有DNA是一半为I5N,另一半是1N的杂合链
图 4.14 密度梯度离心分离 15N DNA 和 14N DNA。(A)离心管的紫外吸收图谱显示两条不 同的 DNA 条带。(B)两条带的紫外吸收峰。 当细菌从 15N-氮源转移到 14N-氮源培养基后,从培养不同时间的细菌抽提 DNA。用密度 梯度离心技术分析。发现繁殖一代的 DNA 样品的位置处于 14N-DNA 和 15N-DNA 条带之间 (图 4.15)。而 15N-DNA 条带(亲本)消失表明 DNA 复制不是全保留的。同样,14N-DNA 条带缺乏表明子代 DNA 有一部分是亲本。由于密度条带刚好是 15N-DNA 条带和 14N-DNA 条带密度和的一半,因此子代 DNA 一半是亲本链,另一半是新生链。 图 4.15 用密度梯度离心检测 E.coli DNA 的半保留复制。DNA 条带的位置取决于 DNA 分子 的 15N 和 14N 含量。复制一代,所有 DNA 是一半为 15N,另一半是 14N 的杂合链