
第二篇 物质代谢及调控 代谢是生命的基本特征之一,借此实现生物体与外界环境的物质交换、自我更新及机体内环境的相对稳定 (图4- 代谢的特点:动态、有序、逐步进行、高度适应和灵敏调节。 新的物质合成 需要能量能 「释放能量谢 目的物质分解 图41代谢示意图 第4章生物氧化 学习要求 了解线粒体外其他氧化体系。 掌握生物氧化的概念,了解生物氧化与燃烧的主 讲义要点 要差异,了 生物氧化 理意义。 (一)生物氧化概述 掌握呼吸链的概念,两条呼吸链的组成成分和排 1,生物氧化的概念物质在生物体内进行的氧 列顺序。掌握氧化磷酸化的概念,熟悉偶联部位,P/0 化反应称为生物氧化,主要是指糖,脂肪、蛋白质等营 比值的概念。了解氧化磷酸化的偶联机制即化学渗 养物质在体内经分解代谢,最终生成C0,和H,0,同 透假说。熟悉影响氧化磷酸化的因素。掌握高能磷 时逐步释放能量,生成ATP供生命活动所需的过程, 酸键的概念熟悉常见的高能陵酸化合物。堂吊ATP 又称细胞氧化或细胞呼吸(表41)。 的生成方式、储存和利用过程 了解胞液中NADH氧 2.氧化反应的主要方式①加氧:②脱氢:③失 化的两种穿梭转运机制。 电子 表41生物氧化与体外氧化的异同 生物氧化 体外氧化 相同点①遵循氧化还原反应的一般规律:有加氧脱氢,失电子等。 ②氧化时的耗氧量,最终的产物,释放的能量均相同。 不同点①反应在有水、体温,近中性的细胞内进行。 ①反应在高温或高压,干燥条件下进行 ③能量逐步释放,部分以化学能方式储存、部分以热能 ③能最问大量经放转拖为光利执 ④产生的C0,和H,0是由物质中的C和H直接与 ④加水脱氢使物质间接获得氧,脱下的氢与氧结合生或水, 氧结合生成 C0,由有机酸脱发产生。 .38
38 第二篇 物质代谢及调控 代谢是生命的基本特征之一ꎬ借此实现生物体与外界环境的物质交换、自我更新及机体内环境的相对稳定 (图 4 ̄1)ꎮ 代谢的特点:动态、有序、逐步进行、高度适应和灵敏调节ꎮ 图 4 ̄1 代谢示意图 第 4 章 生 物 氧 化 学 习 要 求 掌握生物氧化的概念ꎬ了解生物氧化与燃烧的主 要差异ꎬ了解生物氧化的生理意义ꎮ 掌握呼吸链的概念ꎬ两条呼吸链的组成成分和排 列顺序ꎮ 掌握氧化磷酸化的概念ꎬ熟悉偶联部位、P / O 比值的概念ꎮ 了解氧化磷酸化的偶联机制即化学渗 透假说ꎮ 熟悉影响氧化磷酸化的因素ꎮ 掌握高能磷 酸键的概念ꎬ熟悉常见的高能磷酸化合物ꎮ 掌握 ATP 的生成方式、储存和利用过程ꎮ 了解胞液中 NADH 氧 化的两种穿梭转运机制ꎮ 了解线粒体外其他氧化体系ꎮ 讲 义 要 点 (一) 生物氧化概述 1 生物氧化的概念 物质在生物体内进行的氧 化反应称为生物氧化ꎬ 主要是指糖、脂肪、蛋白质等营 养物质在体内经分解代谢ꎬ最终生成 CO2 和 H2Oꎬ同 时逐步释放能量ꎬ生成 ATP 供生命活动所需的过程ꎬ 又称细胞氧化或细胞呼吸(表 4 ̄1)ꎮ 2 氧化反应的主要方式 ①加氧ꎻ②脱氢ꎻ③失 电子ꎮ 表 4 ̄1 生物氧化与体外氧化的异同 生物氧化 体外氧化 相同点 ①遵循氧化还原反应的一般规律:有加氧、脱氢、失电子等ꎮ ②氧化时的耗氧量、最终的产物、释放的能量均相同ꎮ 不同点 ①反应在有水、体温、pH 近中性的细胞内进行ꎮ ②在一系列酶的催化下进行ꎮ ③能量逐步释放ꎬ部分以化学能方式储存、部分以热能 释放ꎮ ④加水脱氢使物质间接获得氧ꎬ脱下的氢与氧结合生成水ꎬ CO2 由有机酸脱羧产生ꎮ ①反应在高温或高压、干燥条件下进行ꎮ ②无须催化或无机催化剂ꎮ ③能量瞬间大量释放ꎬ转换为光和热ꎮ ④产生的 CO2 和 H2O 是由物质中的 C 和 H 直接与 氧结合生成ꎮ

第4章生物氧化·39· (二)生成ATP的氧化体系(线粒体中) ·递氢休和电子传递体的主要功能是传递氢和电子 1.呼吸链 (2H=一2H*+2e). (1)概念:由多种酵和酶构成的递氢体和递电 ·营养物质代谢脱下的氢(NADH和FADH,),通过该 子体按一定顺序排列在线粒体内膜上形成一条使氢 体系的传递,最后与氧结合生成水,同时取动AT甲生成。 ·两条呼吸链:NADH呼吸链和FADH,呼吸继。 氧化成水并释放能量的继锁式反应体系。 (2)呼吸链的组成及排列顺序:见表42、表4.3。 表42线粒体呼吸链成分及主要功能 名称 特点 主要功能 烟酰胺脱氢酶类 以NAD*为铺酶 接受H,传递H(一个H.一个e) 黄素蛋白类 以FAD或FN为辅基 接受H,传递H(2个H) 铁硫蛋白 辅基为铁硫中心或你铁硫能(F。S) 传递单个电子 泛(CaQ) 脂溶性,能在内膜中白由扩散 传H和电子(2个,2个e) 细胞色素类 以血红素为辅基 传逆单个电子 表43线粒体呼吸链复合体 复合体 南名称 亚基 主要成分 知基 作用 复合体1 NADH-CQ还原前 39 黄素蛋白,铁硫蛋白 FMN.Fe-S 将电子从NADH+传递给泛 复合体Ⅱ玻珀酸C0不原酯 4 黄素蛋白、铁硫蛋白 FAD F-S 将中子从玻珀酸传递给衫醒 复合体Ⅲ CoQ-Crte还原酶 10Ct6、Ctcl和铁硫蛋 血红索,S 将电子从还原型泛传递给细胞色煮。 复合体VCtc氧化 13Cta利a3 血红素,C如将电子从细跑色素。传递给氧 以上组分在吸链上的被组装成四个随立的目 表44呼吸徒各传递体的排列顺序 有电子传递能力蛋白质酶复合体。 名称 排列顺序(电位由低到高顺序排列) 但泛醌和C1c均不包含在上述四种复合体中 它们是单独存在的。 NADH氧化呼吸链NADH一→复合体I CQ一复合体Ⅲ Gtc一复合体W一0 (3)呼吸链的电子传递顺序:两条呼吸链的排列 號珀酸氧化呼吸链FAD川,→复合体Ⅱ·CO→复合体Ⅲ 顺序参见表4.4和图4-2。 泛醌(CQ)是两条呼吸链的汇合点。 →Ctc→复合体W→O, 琥珀酸等 复合体I 复合体Ⅲ 复合体N NADH一FMNeS Cyt b -Fe -Cyt e-Cyt c- Cytaa-0 ADP+Pi ATP ADP+Pi ATP ADP+Pi ATP 图4-2呼吸链组成示意图 2.氧化磷酸化 (2)氧化磷酸化的偶联部位:根据氧化还原电势 (1)氧化磷酸化概念:在生物氧化过程中,代谢 与自由能变化关系式,计算出氧化磷酸化的偶联部位 物脱下的氢和电子沿呼吸链传递过程中逐步释放能 存在于复合体I、Ⅲ、V中。因此,在NADH氧化呼吸 量,偶联驱动ADP磷酸化生成ATP。这种氧化与磷酸 链中,有三个部位可以生成AP,而在琥珀酸氧化呼 化紧密偶联的过程即称为氧化磷酸化 吸链中则只有一个部位可以生成ATP」 氧化磷酸化是体内生成ATP的主要方式 (3)P/0比值:P/0比值是指在氧化酶酸化过程
第 4 章 生 物 氧 化 39 (二) 生成 ATP 的氧化体系(线粒体中) 1 呼吸链 (1) 概念:由多种酶和辅酶构成的递氢体和递电 子体按一定顺序排列在线粒体内膜上形成一条使氢 氧化成水并释放能量的链锁式反应体系ꎮ • 递氢体和电子传递体的主要功能是传递氢和电子 (2H 2H + +2e)ꎮ • 营养物质代谢脱下的氢(NADH 和 FADH2 )ꎬ通过该 体系的传递ꎬ最后与氧结合生成水ꎬ同时驱动 ATP 生成ꎮ • 两条呼吸链:NADH 呼吸链和 FADH2 呼吸链ꎮ (2) 呼吸链的组成及排列顺序:见表 4 ̄2、表 4 ̄3ꎮ 表 4 ̄2 线粒体呼吸链成分及主要功能 名称 特点 主要功能 烟酰胺脱氢酶类 以 NAD +为辅酶 接受 Hꎬ传递 H(一个 Hꎬ一个 e) 黄素蛋白类 以 FAD 或 FMN 为辅基 接受 Hꎬ传递 H(2 个 H) 铁硫蛋白 辅基为铁硫中心或称铁硫簇(Fe ̄S) 传递单个电子 泛醌(CoQ) 脂溶性ꎬ能在内膜中自由扩散 传递 H 和电子(2 个 H + ꎬ2 个 e) 细胞色素类 以血红素为辅基 传递单个电子 表 4 ̄3 线粒体呼吸链复合体 复合体 酶名称 亚基 主要成分 辅基 作用 复合体Ⅰ NADH ̄CoQ 还原酶 39 黄素蛋白、铁硫蛋白 FMNꎬFe ̄S 将电子从 NADH+H +传递给泛醌 复合体Ⅱ 琥珀酸 ̄CoQ 还原酶 4 黄素蛋白、铁硫蛋白 FADꎬFe ̄S 将电子从琥珀酸传递给泛醌 复合体Ⅲ CoQ ̄Cyt c 还原酶 10 Cyt b、 Cyt c1 和铁硫蛋白 血红素ꎬFe ̄S 将电子从还原型泛醌传递给细胞色素 c 复合体Ⅳ Cyt c 氧化酶 13 Cyt a 和 a3 血红素ꎬCu 将电子从细胞色素 c 传递给氧 以上组分在呼吸链上的被组装成四个独立的具 有电子传递能力蛋白质 ̄酶复合体ꎮ 但泛醌 和 Cyt c 均不包含在上述四种复合体中ꎬ 它们是单独存在的ꎮ (3) 呼吸链的电子传递顺序:两条呼吸链的排列 顺序参见表 4 ̄ 4 和图 4 ̄2ꎮ 泛醌(CoQ)是两条呼吸链的汇合点ꎮ 表 4 ̄ 4 呼吸链各传递体的排列顺序 名称 排列顺序(电位由低到高顺序排列) NADH 氧化呼吸链 NADH→复合体Ⅰ→ CoQ→复合体Ⅲ → Cyt c→复合体Ⅳ→O2 琥珀酸氧化呼吸链 FADH2→复合体 II→ CoQ→复合体Ⅲ → Cyt c→复合体Ⅳ→O2 图 4 ̄2 呼吸链组成示意图 2 氧化磷酸化 (1) 氧化磷酸化概念:在生物氧化过程中ꎬ代谢 物脱下的氢和电子沿呼吸链传递过程中逐步释放能 量ꎬ偶联驱动 ADP 磷酸化生成 ATPꎮ 这种氧化与磷酸 化紧密偶联的过程即称为氧化磷酸化ꎮ 氧化磷酸化是体内生成 ATP 的主要方式ꎮ (2) 氧化磷酸化的偶联部位:根据氧化 ̄还原电势 与自由能变化关系式ꎬ计算出氧化磷酸化的偶联部位 存在于复合体Ⅰ、Ⅲ、Ⅳ中ꎮ 因此ꎬ在 NADH 氧化呼吸 链中ꎬ有三个部位可以生成 ATPꎬ而在琥珀酸氧化呼 吸链中则只有二个部位可以生成 ATPꎮ (3) P / O 比值:P / O 比值是指在氧化磷酸化过程
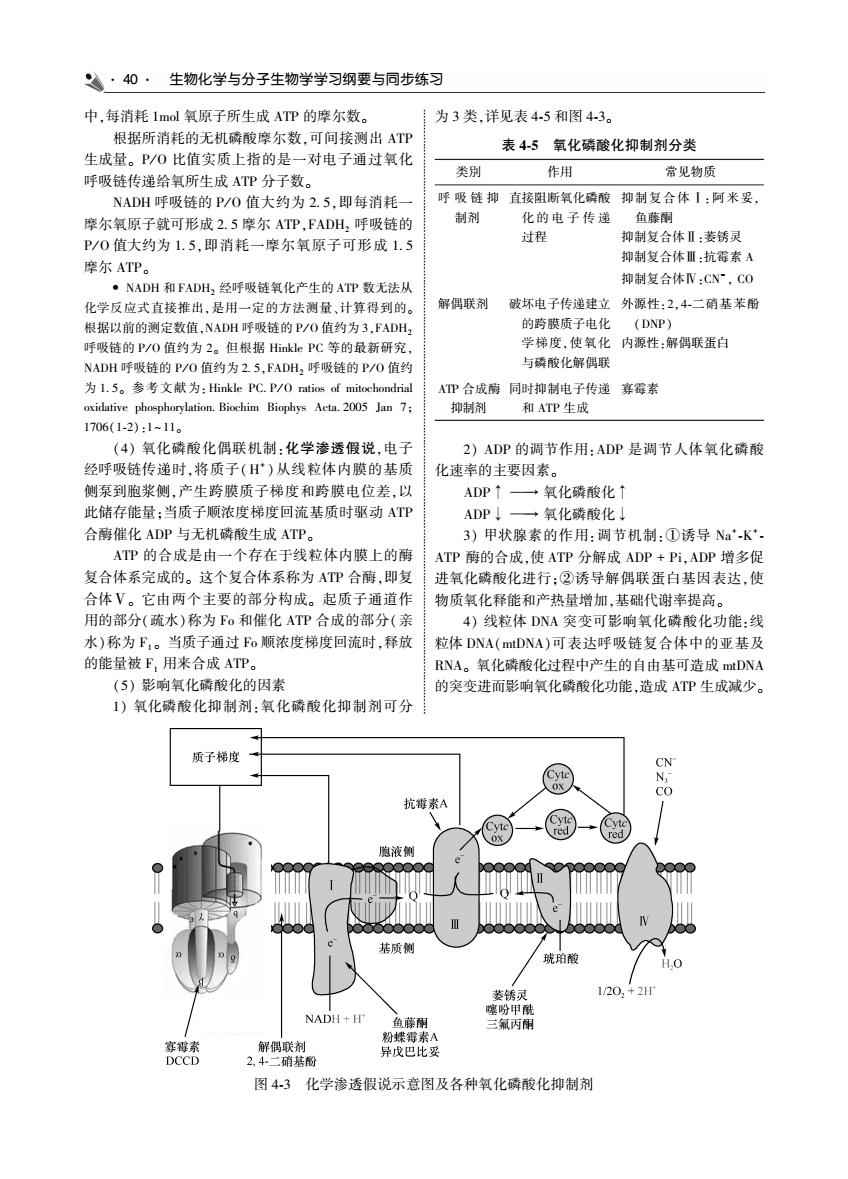
·40·生物化学与分子生物学学习纲要与同步练习 中,每消耗1ml氧原子所生成A的摩尔 为3类,详见表4-5和图4-3。 根据所消耗的 L磷酸摩尔数,可间接测出AT 表45氧化磷酸化抑制剂分类 生成量。P/0比值实质上指的是 一对电子通过氧化 类别 作用 常见物质 呼吸链传递给氧所生成ATP分子数 NADH平吸裤的P/0偵大约为25.即每消耗 呼吸链抑直接阻断氧化磷酸抑制复合体1:阿米妥 摩尔氧原子就可形成2.5摩尔ATP FADH、平吸链的 的电子传递 制 P/0值大约为15即消耗一摩尔氧原子可形成1. 合体Ⅱ:萎锈灵 摩尔ATP 抑制复合体Ⅲ:抗 经呼吸链氧化产生的A 抑制复合体N:CN~,C0C 化学反成式直接据 算得到的 解偶联剂跋坏电子传递建立外源性:2,4.二硝基苯盼 的路随质子由化 (DNP) P/0值约为2。但根我 学梯度使氧化内源性,解偶联蛋自 2.5.FADH 吸链的P 与磷酸化解得联 2057 同时抑制电 传递霉率 和ATP生 1706(1-2).111 (4)氧化磷酸化偶联机制: 化学渗透假说,电了 2)ADP的调节作用:ADP是调节人体氧化磷酸 经呼吸链传递时,将质子()从线粒体内膜的基质 化速率的主要因素。 侧泵到胞浆侧,产生跨膜质子梯度和跨膜电位差,以 ADP T- +氧化磷酸化 此储存能量:当质子顺浓度梯度回流基质时那动AT AD →氧化磷酸化」 合酶催化ADP与无机磷酸生成ATP」 3)用状腺素的作用.调节机制.①诱导N妇K AP的合成是由一个存在于线粒体内膜上的酷 ATP酶的合成,使ATP分解成ADP+Pi,ADP增多促 复合体系完成的。这个复合体系称为ATP合,即复 氧化骑酸化进行:②诱号 解偶联蛋白基因表达,但 合体V。它由两 ~主要的部分构成。起质子通道作 物质氧化释能和产热量增加,基础代谢率提高 用的部分(疏水)称为Fo和催化ATP合成的都分( 4)线粒体DNA突变可影响氧化磷酸化功能:线 水)称为F1。当质子通过o顺浓度梯度回流时,释放 粒体DNA(mDNA)可表达呼吸链复合体中的亚基及 的能量被F,用来合成ATP。 RNA。氧化威酸化过程中产生的自由基可造成tDNA (5)影响氧化磷酸化的因素 的室变讲而影响氧化酸化功能造成ATP生成少 )氧化磷酸化抑制剂:氧化磷酸化抑制剂可分 质子梯度 抗毒素A 一-→ 20 三氣丙雨 解偶剂 二硝 图43化学渗透假说示意图及各种氧化磷酸化抑制剂
40 生物化学与分子生物学学习纲要与同步练习 中ꎬ每消耗 1mol 氧原子所生成 ATP 的摩尔数ꎮ 根据所消耗的无机磷酸摩尔数ꎬ可间接测出 ATP 生成量ꎮ P / O 比值实质上指的是一对电子通过氧化 呼吸链传递给氧所生成 ATP 分子数ꎮ NADH 呼吸链的 P / O 值大约为 2 5ꎬ即每消耗一 摩尔氧原子就可形成 2 5 摩尔 ATPꎬFADH2 呼吸链的 P / O 值大约为 1 5ꎬ即消耗一摩尔氧原子可形成 1 5 摩尔 ATPꎮ • NADH 和 FADH2 经呼吸链氧化产生的 ATP 数无法从 化学反应式直接推出ꎬ是用一定的方法测量、计算得到的ꎮ 根据以前的测定数值ꎬNADH 呼吸链的 P / O 值约为 3ꎬFADH2 呼吸链的 P / O 值约为 2ꎮ 但根据 Hinkle PC 等的最新研究ꎬ NADH 呼吸链的 P / O 值约为 2 5ꎬFADH2 呼吸链的 P / O 值约 为 1 5ꎮ 参考文献为:Hinkle PC P / O ratios of mitochondrial oxidative phosphorylation Biochim Biophys Acta 2005 Jan 7ꎻ 1706(1 ̄2):1~ 11ꎮ (4) 氧化磷酸化偶联机制:化学渗透假说ꎬ电子 经呼吸链传递时ꎬ将质子(H + ) 从线粒体内膜的基质 侧泵到胞浆侧ꎬ产生跨膜质子梯度和跨膜电位差ꎬ以 此储存能量ꎻ当质子顺浓度梯度回流基质时驱动 ATP 合酶催化 ADP 与无机磷酸生成 ATPꎮ ATP 的合成是由一个存在于线粒体内膜上的酶 复合体系完成的ꎮ 这个复合体系称为 ATP 合酶ꎬ即复 合体Ⅴꎮ 它由两个主要的部分构成ꎮ 起质子通道作 用的部分(疏水)称为 Fo 和催化 ATP 合成的部分(亲 水)称为 F1 ꎮ 当质子通过 Fo 顺浓度梯度回流时ꎬ释放 的能量被 F1 用来合成 ATPꎮ (5) 影响氧化磷酸化的因素 1) 氧化磷酸化抑制剂:氧化磷酸化抑制剂可分 为 3 类ꎬ详见表 4 ̄5 和图 4 ̄3ꎮ 表 4 ̄5 氧化磷酸化抑制剂分类 类別 作用 常见物质 呼 吸 链 抑 制剂 直接阻断氧化磷酸 化 的 电 子 传 递 过程 抑制复合体Ⅰ:阿米妥ꎬ 鱼藤酮 抑制复合体Ⅱ:萎锈灵 抑制复合体Ⅲ:抗霉素 A 抑制复合体Ⅳ:CN - ꎬ CO 解偶联剂 破坏电子传递建立 的跨膜质子电化 学梯度ꎬ使氧化 与磷酸化解偶联 外源性:2ꎬ4 ̄二硝基苯酚 (DNP) 内源性:解偶联蛋白 ATP 合成酶 抑制剂 同时抑制电子传递 和 ATP 生成 寡霉素 2) ADP 的调节作用:ADP 是调节人体氧化磷酸 化速率的主要因素ꎮ ADP↑ → 氧化磷酸化↑ ADP↓ → 氧化磷酸化↓ 3) 甲状腺素的作用:调节机制:①诱导 Na +  ̄K +  ̄ ATP 酶的合成ꎬ使 ATP 分解成 ADP + PiꎬADP 增多促 进氧化磷酸化进行ꎻ②诱导解偶联蛋白基因表达ꎬ使 物质氧化释能和产热量增加ꎬ基础代谢率提高ꎮ 4) 线粒体 DNA 突变可影响氧化磷酸化功能:线 粒体 DNA(mtDNA)可表达呼吸链复合体中的亚基及 RNAꎮ 氧化磷酸化过程中产生的自由基可造成 mtDNA 的突变进而影响氧化磷酸化功能ꎬ造成 ATP 生成减少ꎮ 图 4 ̄3 化学渗透假说示意图及各种氧化磷酸化抑制剂
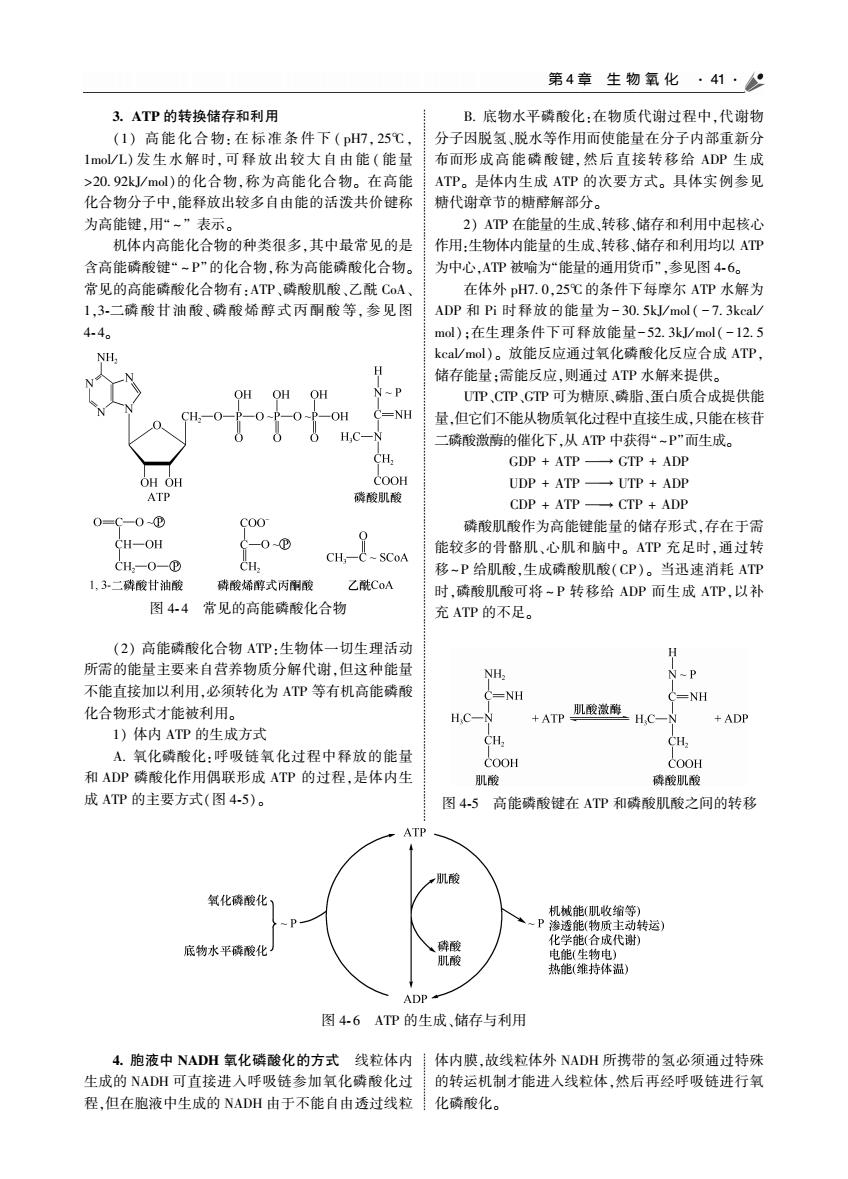
第4章生物氧化·41·9 3.ATP的转换储存和利用 B.底物水平磷酸化:在物质代谢过程中,代谢物 (1)高能化合物.在标准条件下(DH7.25℃ 分子因脱氢,脱水等作用而使能量在分子内部重新分 1mo/L)发生水解时,可释放出较大自由能(能量 布而形成高能磷酸键,然后直接转移给ADP生成 >20.92J/ml)的化合物,称为高能化合物。在高能 ATP。是体内生成ATP的次要方式。具体实例参见 化合物分子中,能释放出较多自由能的 代谢章节的糖辞解部分 为高 e ATP在能量的生成转移,储存和利用中起核心 机体内高能化合物的种类很多,其中最常见的是 作用:生物体内能量的生成,转移,储存和利用均以ATP 含高能磷酸键“-P“的化合物,称为高能磷酸化合物。 为中心,AP被哈为“能量的通用货币”,参见图4-6。 常见的高能磷酸化合物有:ATP,磷酸肌酸、乙酰CA,」 在体外DH7.0.25℃的条件下每摩尔ATP水解为 13.一硫酸甘油酸、磷酸烯醇式丙酮酸等,参见图 ADP和Pi时释放的能量为-30.5J/mol(-7.3keal 44。 mal):在生理条件下可释放能量-52.3/ml(-12.5 NH. 放能反应通过氧化磷酸化反应合成ATP 储存能量:需能反应,则通过ATP水解来提供。 OH OH UTP,CIPGTP可为糖原、磷脂、蛋白质合成提供横 量,但它们不能从物质氧化过程中直接生成,只能在核苷 H.O 二磷酸微剧的催化下,从AP中获得“-P”面生成。 CH. CDD TDCTDADD OOH UDP+ATP -UTP +ADP ATP 磷酸肌酸 CTP 磷酸肌酸作为高能键能量的储存形式,存在于需 能较多的骨酪肌、心肌和脑中。ATP充足时,通过转 CH,-( SCoA 移-P给肌酸,生成磷酸肌酸(CP)。当迅速消耗ATP 二磷酸甘油酸 酸烯醇式丙酮酸 时磷酸肌酸可将~P转移给ADP而生成AP以补 图44常见的高能醉酸化合物 充ATP的不足 (2)高能磷酸化合物ATP:生物体一切生理活动 H 所需的能量主要来自营养物质分解代谢,但这种能量 不能直接加以利用,必须转化为AP等右机高能威酸 -NH NH 化合物形式才能被利用。 HC-N +AP肌酸激影 H.C-N +ADP 休内 ATP的生成方式 CH. A氧化醉酸化:呼吸链氧化过程中释放的能到 和ADP磷酸化作用偶联形成ATP的过程,是体内生 肌酸 破酸肌酸 成ATP的主要方式(图4-5)。 图4-5高能磷酸键在ATP和磷酸肌酸之间的转移 氧化障酸化 机械能(肌收缩等 转运 成代谢 底物水平碎酸化 热能(维持体温 ADP 图4-6ATP的生成、储存与利用 4.胞液中NADH氧化磷酸化的方式线粒体内!体内膜,故线粒体外NADH所携带的氢必须通过特殊 生成的NADH可直接进入呼吸链参加氧化磷酸化过的转运机制才能进人线粒体,然后再经呼吸链进行氧 程但在胞液中生成的NADH由于不能自由诱过线粒 化酸化
第 4 章 生 物 氧 化 41 3 ATP 的转换储存和利用 (1) 高能化合物: 在标准条件下 ( pH7ꎬ 25℃ ꎬ 1mol / L) 发生水解时ꎬ可释放出较大自由能 ( 能量 >20 92kJ/ mol)的化合物ꎬ称为高能化合物ꎮ 在高能 化合物分子中ꎬ能释放出较多自由能的活泼共价键称 为高能键ꎬ用“ ~ ” 表示ꎮ 机体内高能化合物的种类很多ꎬ其中最常见的是 含高能磷酸键“ ~ P”的化合物ꎬ称为高能磷酸化合物ꎮ 常见的高能磷酸化合物有:ATP、磷酸肌酸、乙酰 CoA、 1ꎬ3 ̄二磷酸甘油酸、磷酸烯醇式丙酮酸等ꎬ参见图 4 ̄ 4ꎮ 图 4 ̄ 4 常见的高能磷酸化合物 (2) 高能磷酸化合物 ATP:生物体一切生理活动 所需的能量主要来自营养物质分解代谢ꎬ但这种能量 不能直接加以利用ꎬ必须转化为 ATP 等有机高能磷酸 化合物形式才能被利用ꎮ 1) 体内 ATP 的生成方式 A 氧化磷酸化:呼吸链氧化过程中释放的能量 和 ADP 磷酸化作用偶联形成 ATP 的过程ꎬ是体内生 成 ATP 的主要方式(图 4 ̄5)ꎮ B 底物水平磷酸化:在物质代谢过程中ꎬ代谢物 分子因脱氢、脱水等作用而使能量在分子内部重新分 布而形成高能磷酸键ꎬ然后直接转移给 ADP 生成 ATPꎮ 是体内生成 ATP 的次要方式ꎮ 具体实例参见 糖代谢章节的糖酵解部分ꎮ 2) ATP 在能量的生成、转移、储存和利用中起核心 作用:生物体内能量的生成、转移、储存和利用均以 ATP 为中心ꎬATP 被喻为“能量的通用货币”ꎬ参见图 4 ̄ 6ꎮ 在体外 pH7 0ꎬ25℃ 的条件下每摩尔 ATP 水解为 ADP 和 Pi 时释放的能量为- 30 5kJ/ mol ( - 7 3kcal / mol)ꎻ在生理条件下可释放能量-52 3kJ/ mol( -12 5 kcal/ mol)ꎮ 放能反应通过氧化磷酸化反应合成 ATPꎬ 储存能量ꎻ需能反应ꎬ则通过 ATP 水解来提供ꎮ UTP、CTP、GTP 可为糖原、磷脂、蛋白质合成提供能 量ꎬ但它们不能从物质氧化过程中直接生成ꎬ只能在核苷 二磷酸激酶的催化下ꎬ从 ATP 中获得“ ~P”而生成ꎮ GDP + ATP → GTP + ADP UDP + ATP → UTP + ADP CDP + ATP → CTP + ADP 磷酸肌酸作为高能键能量的储存形式ꎬ存在于需 能较多的骨骼肌、心肌和脑中ꎮ ATP 充足时ꎬ通过转 移~ P 给肌酸ꎬ生成磷酸肌酸(CP)ꎮ 当迅速消耗 ATP 时ꎬ磷酸肌酸可将 ~ P 转移给 ADP 而生成 ATPꎬ以补 充 ATP 的不足ꎮ 图 4 ̄5 高能磷酸键在 ATP 和磷酸肌酸之间的转移 图 4 ̄ 6 ATP 的生成、储存与利用 4 胞液中 NADH 氧化磷酸化的方式 线粒体内 生成的 NADH 可直接进入呼吸链参加氧化磷酸化过 程ꎬ但在胞液中生成的 NADH 由于不能自由透过线粒 体内膜ꎬ故线粒体外 NADH 所携带的氢必须通过特殊 的转运机制才能进入线粒体ꎬ然后再经呼吸链进行氧 化磷酸化ꎮ
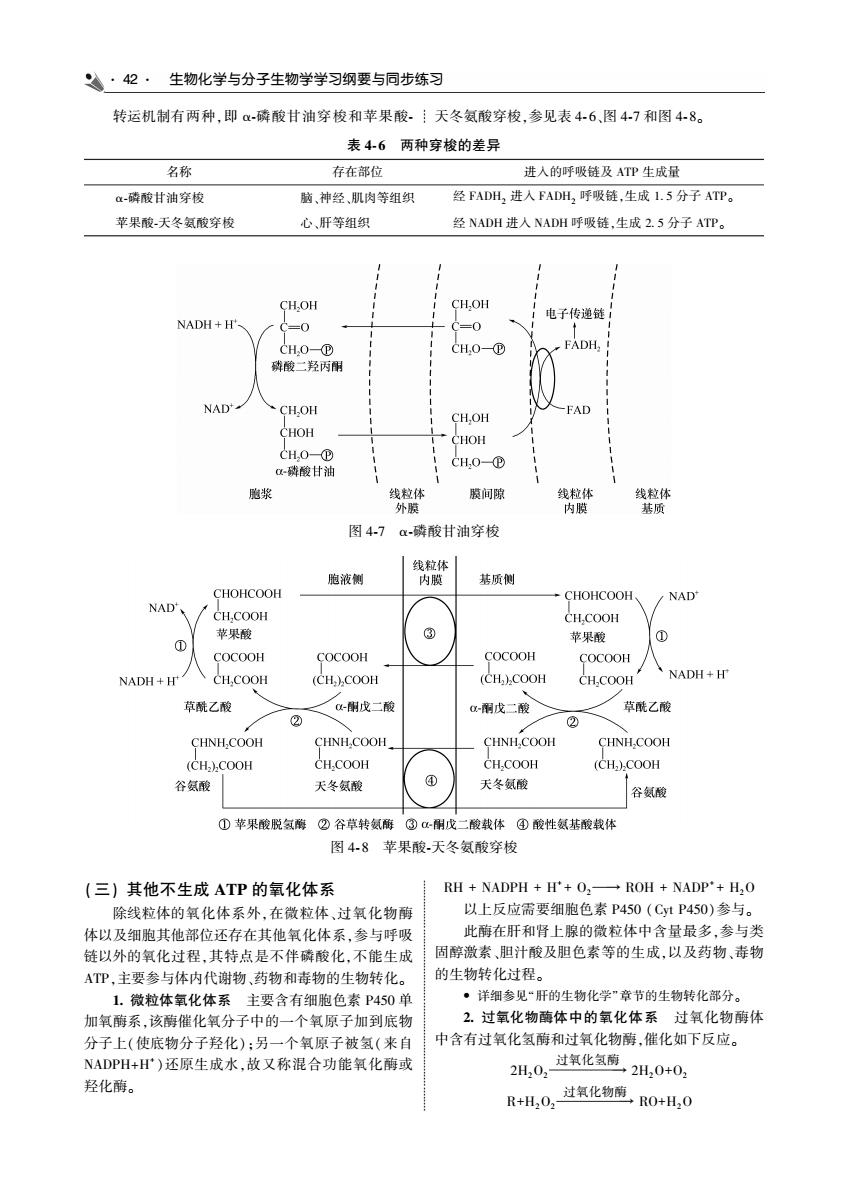
·42·生物化学与分子生物学学习纲要与同步练习 转运机制有两种,即α磷酸甘油穿梭和苹果酸 天冬氨酸穿梭,参见表46,图4-7和图48 表46两种穿梭的差异 名称 存在部位 进入的呼吸皓及ATP生成量 酶酸甘油穿 脑、神经、肌肉等组织 经FADH,进人FADH,吸链,生成1.5分子ATP。 苹果酸天冬氨酸穿梭 广肝等组组 经NADH进人NADH呼吸链,生成2.5分子ATP。 CH.OH 电子传递链 NADH+H CH0- FADH, NAD CH.OH CH.OH FAD CHOH CHOH CH,0-⑧ 胞浆 膜间隙 找 图47a磷酸甘油穿梭 胞液侧 基质侧 CHOHCOOH CHOHCOOH NAD NAD /CH.COOH 苹果酸 ③ COCOOH NADH+H CH.COOH (CH).COOH (CH.).COOH CH.COOH 丶NADH+F 草酰乙酸 、戊二酸 酮戊二酸 点能乙酸 ② CHNH COOH CHNH.COOH CHNH,COOH CHNHCOOH CH.)COOH CH.COOH CH.COOH (CH.)COOH 容氨酸 天冬氨酸 天冬氨酸 容氨酸 ①苹果酸脱氢酶②谷草转氨稀③α:刚戊二酸载体④酸性氨基酸载体 图48苹果酸天冬氨酸穿梭 ((三)其他不生成ATP的氧化体系 RH NADPH+H'+O. ROH NADP+H,O 除线粒体的氧化体系外,在微粒体、过氧化物酶 以上反应需要细胞色素P450(C1P 50)参与 体以及细胞其他部位还存在其他氧化体系,参与呼吸 此酶在肝和肾上腺的微粒体中含量最多,参与类 链以外的氧化过程,其特点是不伴磷酸化,不能生成 固醇激素、胆汁酸及胆色素等的生成,以及药物、毒物 ATP,主要参与体内代谢物、药物和毒物的生物转化。 的生物转化时程。 1.微粒体氧化体 主要含有细 胞色素 ·详细参见“肝的生物化学”章节的生物转化部分】 FP430 加氧酶系,该酶催化氧分子中的一个氧原子加到底物 2.过氧化物酶体中的氧化体系过氧化物酶体 分子上(使底物分子羟化):另一个氧原子被氢(米自 中含有过氧化氢酶和过氧化物爵,催化如下反应 NADPH+Hr)还原生成水,故又称混合功能氧化南或 20,过氧化氢2H,0+0, 羟化酶。 R+H0,过氧化商RO+H.0
42 生物化学与分子生物学学习纲要与同步练习 转运机制有两种ꎬ即 α ̄磷酸甘油穿梭和苹果酸 ̄ 天冬氨酸穿梭ꎬ参见表 4 ̄ 6、图 4 ̄7 和图 4 ̄ 8ꎮ 表 4 ̄ 6 两种穿梭的差异 名称 存在部位 进入的呼吸链及 ATP 生成量 α ̄磷酸甘油穿梭 脑、神经、肌肉等组织 经 FADH2 进入 FADH2 呼吸链ꎬ生成 1 5 分子 ATPꎮ 苹果酸 ̄天冬氨酸穿梭 心、肝等组织 经 NADH 进入 NADH 呼吸链ꎬ生成 2 5 分子 ATPꎮ 图 4 ̄7 α ̄磷酸甘油穿梭 图 4 ̄ 8 苹果酸 ̄天冬氨酸穿梭 (三) 其他不生成 ATP 的氧化体系 除线粒体的氧化体系外ꎬ在微粒体、过氧化物酶 体以及细胞其他部位还存在其他氧化体系ꎬ参与呼吸 链以外的氧化过程ꎬ其特点是不伴磷酸化ꎬ不能生成 ATPꎬ主要参与体内代谢物、药物和毒物的生物转化ꎮ 1 微粒体氧化体系 主要含有细胞色素 P450 单 加氧酶系ꎬ该酶催化氧分子中的一个氧原子加到底物 分子上(使底物分子羟化)ꎻ另一个氧原子被氢(来自 NADPH+H + )还原生成水ꎬ故又称混合功能氧化酶或 羟化酶ꎮ RH + NADPH + H + + O2 → ROH + NADP + + H2O 以上反应需要细胞色素 P450 (Cyt P450)参与ꎮ 此酶在肝和肾上腺的微粒体中含量最多ꎬ参与类 固醇激素、胆汁酸及胆色素等的生成ꎬ以及药物、毒物 的生物转化过程ꎮ • 详细参见“肝的生物化学”章节的生物转化部分ꎮ 2 过氧化物酶体中的氧化体系 过氧化物酶体 中含有过氧化氢酶和过氧化物酶ꎬ催化如下反应ꎮ 2H2O2 过氧化氢酶 → 2H2O+O2 R+H2O2 过氧化物酶 → RO+H2O