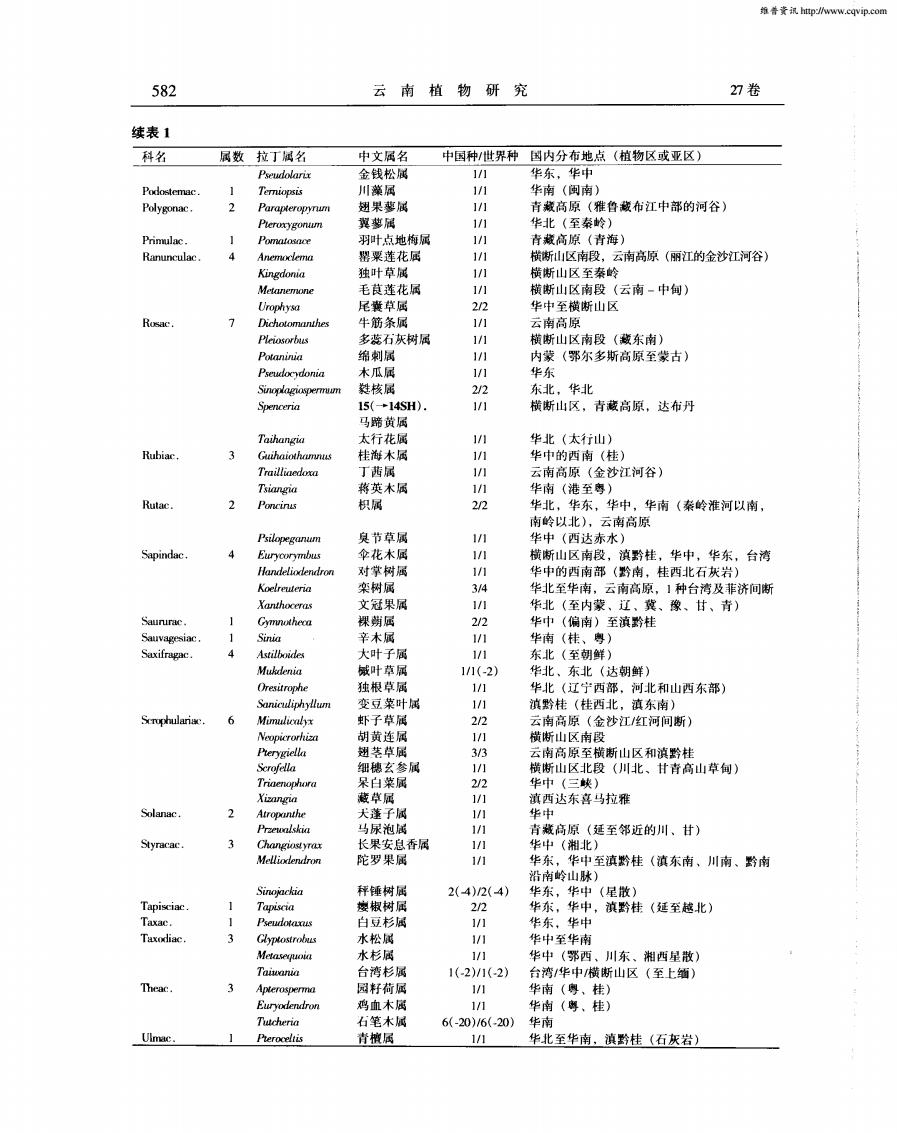
维鲁资讯http://www.cqvip.com27卷582云南植物研究续表1科名中国种/世界种国内分布地点(植物区或亚区)属数拉丁属名中文属名华东,华中金钱松属1/1Pseudolarir川藻属1/1华南(闽南)Podostemac.1Termiopsis2翅果薯属1/1青藏高原(雅台藏布江中部的河谷)Polygonac.Parapteropyram翼萝属1/1华北(至秦岭)Pleroxygonun羽叶点地梅属1/1青激高原(青海)Primulac.Pomatosnce1Ranunculac.4墨栗莲花属1/1Anemoclema横断山区南段,云南高原(丽江的金沙江河谷)独叶草属1/1横断山区至秦岭Kingdonia1/1Metanemone毛莲花属横断山区南段(云南-中甸)尾囊草属2/2华中至横断山区UrophysaRosac:牛筋条属1/17Dichotomanthes云南高原Pleiosorbes多蕊石灰树属1/1横断山区南段(藏东南)1/1Potaninia绵刺属内蒙(鄂尔多斯高原至蒙古)木瓜属1/1华东Pseudocydonia2/2东北,华北核属Sinoplagiospermuam1/115(-14SH).横断山区,青藏高原,达布丹Spenceria马蹄黄属Taihangia太行花属1/1华北(太行山)1/1Rubiac.3Guihaiothamus桂海木属华中的西南(桂)Trailliaedoxa丁茜属1/1云南高原(金沙江河谷)蒋英木属1/1Tsiangia华南(港至粤)2积属212Rutac.Poncinus华北,华东,华中,华南(秦岭淮河以南,南岭以北),云南高原Psilopeganum臭节草属1/1华中(西达赤水)1/1Sapindac.4伞花木属横断山区南段,滇黔桂,华中,华东,台湾EurycorymbusHandeliodendron对掌树属1/1华中的西南部(黔南,桂西北石灰岩)栾树属3/4华北至华南,云南高原,1种台湾及菲济间断Koelreuteria文冠果属1/1Xanthoceras华北(至内蒙、辽、冀、豫、甘、青)裸剪属2/2Saunurac.1Cymnotheca华中(偏南)至滇黔桂Sauvagesiac.1辛木属1/1Sinia华南(桂、粤)4大叶子属1/1Saxifragac.Astilboides东北(至朝鲜)叶草属华北、东北(达朝鲜)Mukdenia1/1(-2)独根草属1/1Oresitrophe华北(辽宁西部,河北和山西东部)变豆菜叶属1/1Sanicuiphyllum滇黔桂(桂西北,滇东南)虾子草属2/2Serophulariac.6Mimuliealyx云南高原(金沙江/红河间断)胡黄连属1/1Neopicrorhiza横断山区南段翅芒草属3/3Prerygiella云南高原至横断山区和滇黔桂1/1Scrofella细穗玄参属横断山区北段(川北、甘青高山草甸)Triaenophara呆白菜属2/2华中(三峡)藏草属1/1Xizangia滇西达东喜马拉雅Solanac.2犬莲子属1/1华中AtropanthePrzewalskia马尿泡属1/1青藏高原(延至邻近的川、甘)3长果安息香属1/1Styracac.Changiastyrax华中(湘北)陀罗果属1/1Melliodendron华东,华中至滇黔桂(滇东南、川南、黔南沿南岭山脉)Sinojackia秤锤树属2(-4)/2(-4)华东,华中(星散)Tapisciac.1Tapiscia椒树属2/2华东,华中,滇黔桂(延至越北)Taxac.11/1华东,华中Pseudotaxus白豆杉属Taxodiac.3水松属1/1Glyptostrobus华中至华南水杉属1/1华中(鄂西、川东、湘西星散)MetasequoiaTaiwania台湾杉属1(-2)/1(-2)台湾/华中/横断山区(至上缅)Theac.31/1园籽荷属华南 (粤、桂)Apterospermia1/1Euryodendron鸡血木属华南 (粤、桂)Tucheria石笔木属6(-20)/6(-20)华南Ulmac青属1/1Preroceltis华北至华南,滇黔桂(石灰岩)
582 云 南 植 物 研 究 27卷 维普资讯 http://www.cqvip.com
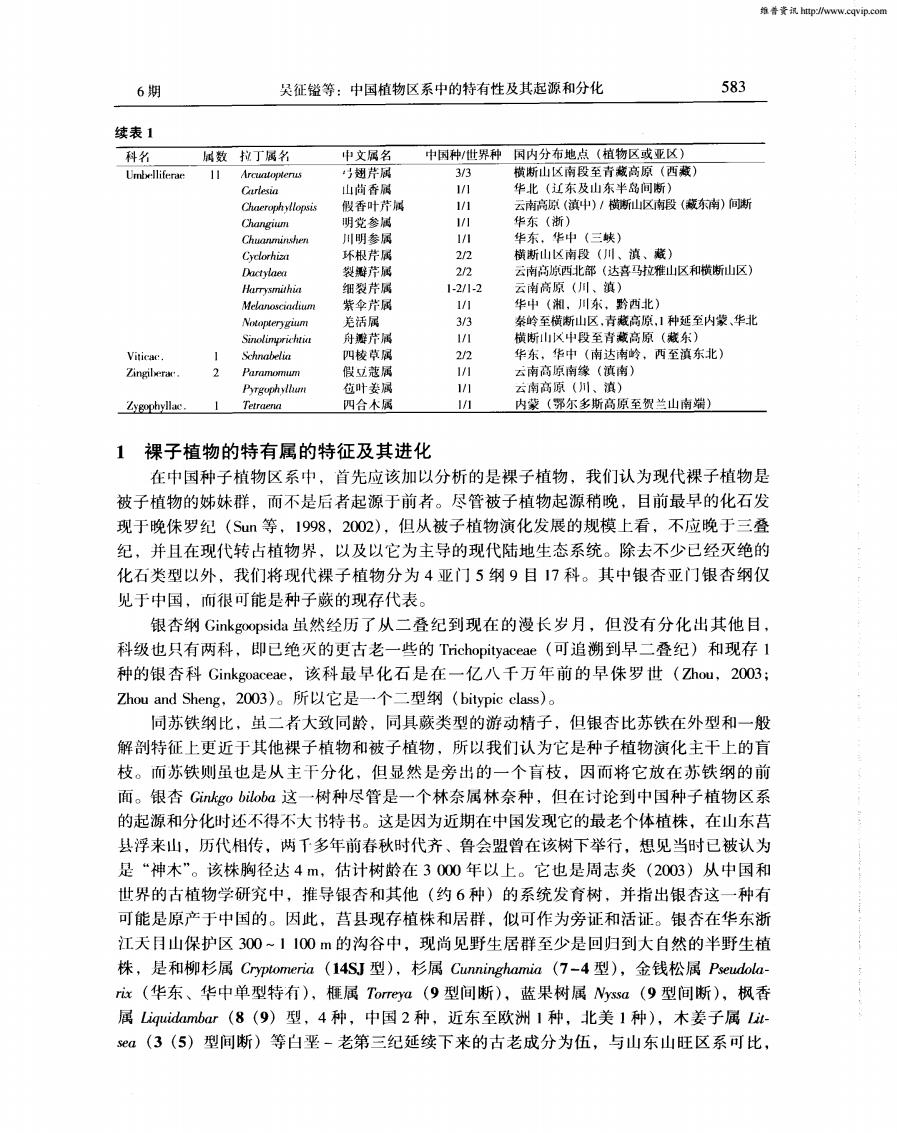
维鲁资http://www.cqvip.com583吴征镒等:中国植物区系中的特有性及其起源和分化6期续表1国内分布地点(植物区或亚区)科名属数拉丁属名中文属名中国种/世界种3/3横断山区南段至青藏高原(西藏)巧翅芹属UmbellifenareArcuntoptenus111/1华北(辽东及山东半岛间断)Curlesia山香属1/1云南高原(滇中)/横断山区南段(藏东南)向断假香叶芹属Chaerophytlopsis1/1华东 (浙)Changiun明党参属1/1华东,华中(三峡)川明参属Chuanminshen2/2横断山区南段(川、滇、葳)环根芹属Cyclorhiza2/2裂摄芹属云南高原西北部(达喜马拉雅山区和横断山区)Dactylaea细裂芹属1-2/1-2云南高原(川、滇)Harrysmithia紫伞芹属1/1华中(湘,川东,黔西北)Melanosciadlium3/3秦岭至横断山区,青藏高原,1种延至内蒙、华北Notopterygium羌活属1/1横断山区中段至青藏高原(藏东)舟墨芹属Sinolimprichtiar2/2Schnabelia四棱草属华东、华中(南达南岭、西至滇东北)Viticac.11/1假立蔻属云南高原南缘(滇南)Zingilerac2Paramomum1/1Pyrgophyluan苞叶姜属云南高原(川、滇)11Tetraena四合木属内蒙(鄂尔多斯高原至贺兰山南端)Zygophyllac裸子植物的特有属的特征及其进化1在中国种子植物区系中,首先应该加以分析的是裸子植物,我们认为现代裸子植物是被子植物的姊妹群,而不是后者起源于前者。尽管被子植物起源稍晚,目前最早的化石发现于晚侏罗纪(Sun等,1998,2002),但从被子植物演化发展的规模上看,不应晚于三登纪,并且在现代转占植物界,以及以它为主导的现代陆地生态系统。除去不少已经灭绝的化石类型以外,我们将现代裸子植物分为4亚门5纲9目17科。其中银杏亚门银杏纲仅见于中国,而很可能是种子蕨的现存代表。银杏纲Ginkgoopsida虽然经历了从二叠纪到现在的漫长岁月,但没有分化出其他目,科级也只有两科,即已绝灭的更古老一些的Trichopityaceae(可追溯到早二叠纪)和现存1种的银杏科Ginkgoaceae,该科最早化石是在一亿八千万年前的早侏罗世(Zhou,2003;ZhouandSheng,2003)。所以它是一个二型纲(bitypicclass)。同苏铁纲比,虽二者大致同龄,同具蕨类型的游动精子,但银比苏铁在外型和一般解剖特征上更近于其他裸子植物和被子植物,所以我们认为它是种子植物演化主干上的盲枝。而苏铁则虽也是从主干分化,但显然是旁出的一个盲枝,因而将它放在苏铁纲的前面。银杏Ginkgobiloba这一树种尽管是一个林奈属林奈种,但在讨论到中国种子植物区系的起源和分化时还不得不大书特书。这是因为近期在中国发现它的最老个体植株,在山东营县浮来山,历代相传,两千多年前春秋时代齐、鲁会盟曾在该树下举行,想见当时已被认为是“神木”。该株胸径达4m,估计树龄在3000年以上。它也是周志炎(2003)从中国和世界的古植物学研究中,推导银杏和其他(约6种)的系统发育树,并指出银杏这一种有可能是原产于中国的。因此,营县现存植株和居群,似可作为旁证和活证。银查在华东浙江天日山保护区300~1100m的沟谷中,现尚见野生居群至少是回归到大自然的半野生植株,是和柳杉属Cryptomeria(14SJ型),杉属Cunninghamia(7-4型),金钱松属Pseudola-rix(华东、华中单型特有)、属Torreya(9型间断),蓝果树属Nyssa(9型间断),枫香属Liquidambar(8(9)型,4种,中国2种,近东至欧洲1种,北美1种),木姜子属Lit-sea(3(5)型间断)等白垩-老第三纪延续下来的古老成分为伍,与山东山旺区系可比
6期 吴征镒等 :中国植物区系中的特有性 及其起源和分化 583 l 裸 子植物 的特有属 的特征及 其进化 在中国种子植物 区系巾,首先应该加以分析的是裸子植物 ,我们认为现代裸子植物是 被子植物的姊妹群 ,而不是后者起源于前者。尽管被子植物起源稍晚 ,目前最早的化石发 现于晚侏罗纪 (Sun等,l998,2002),但从被子植物演化发展的规模上看 ,不应晚于三叠 纪 ,并且在现代转 占植物界 ,以及以它为主导的现代陆地生态系统 。除去不少已经灭绝 的 化石类型以外 ,我们将现代裸 子植物分为 4亚门 5纲 9目 l7科 。其中银杏亚门银杏纲仅 见于中国 ,而很可能是种子蕨的现存代表。 银杏纲 Ginkgoopsida虽然经历 了从二叠纪到现在的漫长岁月 ,但没有分化出其他 目, 科级也只有两科 ,即已绝灭的更古老一些的 Trichopityaceae(可追溯到早二叠纪)和现存 1 种的银杏科 Ginkgoaceae,该科 最早化石 是在一亿 八千万 年前 的早侏 罗世 (Zhou,2003; ZhouandSheng,2003)。所 以它 是一个 二 型纲 (bitypicclass)。 同苏铁纲比,虽二者大致同龄 ,同具蕨类型的游动精子 ,但银杏 比苏铁在外型和一般 解剖特征上更近于其他裸子植物和被子植物 ,所以我们认 为它是种子植物演化主干上的盲 枝 。而苏铁则虽也是从主 F分化 ,但显然是旁出的一个盲枝 ,因而将它放在苏铁 纲的前 而。银杏 Ginkgobiloba这一树种尽管是一个林奈属林奈种 ,但在讨论到中国种子植物 区系 的起源和分化时还不得不大_f5特书。这是因为近期在中国发现它的最老个体植株,在 山东莒 县浮来山,历代相传,两 T多年前春秋时代齐 、鲁会盟曾在该树下举行,想见当时 已被认为 足 “神木”。该株胸径达 4m,估计树龄在 3000年 以上。它也是周志炎 (2003)从 中国和 世界的古植物学研究中 ,推导银杏和其他 (约 6种)的系统发育树 ,并指出银杏这一种有 可能是原产于中国的。因此,莒县现存植株和居群,似可作为旁证和活证。银杏在华东浙 江天 日山保护区 300~l100m的沟谷中,现尚见野生居群至少是 回归到大自然的半野生植 株 ,是和柳 杉属 Cryptomer/a (14S,I型 ),杉属 Cunninghamia (7—4型 ),金钱 松属 Pseudola— fix (华东 、华中单型特有),榧属 Torreya (9型间断),蓝果树属 Nyssa (9型间断 ),枫香 属 Liquidambar(8(9)型 ,4种 ,中国 2种 ,近东至欧洲 1种 ,北美 1种 ),木姜子属 一 sea (3(5)型间断)等白垩 一老第三纪延续下来的古老成分为伍 ,与 山东山旺区系可比, 维普资讯 http://www.cqvip.com

维普资in.htp://www.cqvip.com27卷584云南植物研究组成中生混交林的。银杏在中国的文献记载虽然只从唐代开始,在段成式的《西阳杂短》中,对这一树种的丛生而怀中孢子的现象有所描述,想来他是从四川酉、秀、黔、彭这一带的“水杉区系”(Hu,1980)中,见到过野生植株。它当时广泛出现在华中类型的森林中,往后向应海也在黔东北见到此种现象,哈佛大学阿诺德树木园银杏专家PelerTredici博土实地对黔东北野生银查进行了考察,确信为野生的银杏群落(孙航同PeterTredici私人通信)。我国自古以来银杏便广为栽培和利用,在道教和佛教寺观和孔夫子文庙中常多古代栽培或开山时保留下来的古树,迄今在滇东北的保护林中也往往见到古老的大根盘(孙必兴告知)似乎应该是滇东北经黔东北至华北(东北)至华中,华东是它的原产地。它在现代中国植物区系中4个特有单型科的首位,而县的植株无疑是中国原产的“世界爷”,是比北美加州所产Sequciodendrongiganteum年龄可能还要古老的物种的个体。苏铁门亦仅存苏铁纲Cycadopsida,但Boweniaceae,Stangeriaceae和Zamiaceae3个稍大科不见于中国,中国也没有特有属。狭义的苏铁科仅苏铁(Cycas)1属,主要分布于旧世界热带和亚热带地区。该属目前分为三亚属即:苏铁亚属(主要分布于中国和中南半岛),拳叶苏铁亚属(主要分布于中南半岛至印度),刺叶苏铁亚属(主要分布于澳大利亚及其附近岛屿),世界有86种,中国有22~25种(或至少16种),且有较原始的林下种类“Epicycas”。(Wang,1996,2001;Delaubenfels andAdema,1998),关于苏铁的起源时间,我国学者朱家等有较深入的研究,朱家、杜贤铭(1981)在山西太原二登世地层发现了中国始苏铁(Primocycas chinensis),其掌状深裂的大孢子叶与现代苏铁属十分相似,从而将苏铁起源的时间推到了二叠纪。朱家等(1994)又进一步发现了苏铁小孢子叶球古生铁花(Cycadostrobiluspaleozoicus),证明了我国华北地区早在古生代二叠纪就有苏铁的生长。最近,胡雨帆等(Hu等,1999)根据采自甘肃石炭世苏铁类大孢子叶化石,推论苏铁类植物很可能起源于我国西北地区。管中天(1996)并认为在古生代华北地区是苏铁类植物的分布中心,但在中生代则移向西南地区。由于西南地区自古生代以来地质环境和气候条件一直相对较稳定,可为苏铁植物的进一步发生发展提供良好的条件。此外,较为丰富的苏铁和种子蕨化石的记录(徐仁等,1979)为此提供了古植物学的证据。从苏铁属的现代地理分布上分析,根据王定跃(Wang,2001)的研究,虽然现代苏铁属的二大分布中心在中国西南和澳大利亚及其邻近岛屿,但我国西南地区至中南半岛地区是该属原始类群苏铁亚属的集中分布区域,同时也包含了不同进化水平的3个亚属的类群,因此,我国的西南地区很可能是现代苏铁的发源地。松柏亚门有二纲,松柏纲含6科,我国缺南洋杉科(Araucariaceae),贝壳杉科(Agathidaceae),金松科(Sciadopityaceae)3科。尽管金松科见于日本中部至南部,不达我国。但它比中国出现的柏、杉、松3科为古老。柏科(Cupressaceae)是从三叠纪到早侏罗纪起源和分化的两温带分布大科。在1820属中,北温带分布即有13属,而我国有其11,东亚-北美分布型2个,北温带分布型3个,东亚分布型4个。故而科的分布型用复合表示,应为南北温带广布,即8-4(9,14)型。其起源应该认为是在泛古大陆时,古北大陆东部(E。Laurasia),是逐渐向古南大陆西部扩散的。我国该科虽然没有一个特有属,却有2个准特有属,1是翠柏属(Calo
584 云 南 植 物 研 究 27卷 组 成 中生混 交林 的 。 银杏在中国的文献记载虽然只从唐代开始 ,在段成式的 《酉阳杂俎》中,对这一树种 的丛生而怀中孢子 的现象有所描述 ,想来他是从 四川酉 、秀 、黔 、彭这一带 的 “水杉 区 系”(Hu,1980)中,见到过野生植株。它 当时广泛 出现在华 中类型的森林中,往后 向应 海也在黔东北见到此种现象 ,哈佛大学阿诺德树木园银 杏专 家 PeterTredici博 士实地对黔 东北野生银杏进行 了考察 ,确信为野生的银杏群落 (孙航 同 PeterTredici私人通信)。我 国 自古以来银杏便广为栽培和利用 ,在道教和佛教寺观和孔夫子文庙 中常多古代栽培或开山 时保留下来的古树 ,迄今在滇东北的保护林 中也往往见到古老 的大根盘 (孙必兴告知), 似乎应该是滇东北经黔东北至华北 (东北 )至华中,华东是它的原产地 。它在现代中国植 物区系中 4个特有单型科的首位,而莒县的植株无疑是 中国原产 的 “世界爷”,是 比北美 加州所产 Sequciodendrongiganteum年龄可能还要古老的物种的个体 。 苏铁 门亦仅 存 苏 铁 纲 Cyeadopsida,但 Boweniaceae,Stangeriaceae和 Zamiaceae3个 稍 大 科不见于中国,中国也没有特有属。狭义的苏铁科仅苏铁 (Cycas)1属 ,主要分布于旧世 界热带和亚热带地 区。该属 目前分为三亚属即:苏铁亚属 (主要分布于中国和中南半岛), 拳叶苏铁亚属 (主要分布于中南半岛至印度),刺 叶苏铁亚属 (主要分布于澳大利亚及其 附近岛屿),世界有 86种,中国有 22~25种 (或 至少 l6种 ),且有较原始 的林 下种类 “ 咖 ”。 (Wang,1996,2001;DelaubenfelsandAdema,1998),关 于苏 铁 的 起 源 时 间 , 我 国学者朱家槽等有较深入 的研究 ,朱家槽 、杜贤铭 (1981)在山西太原二叠世地层发现 了中国始苏铁 (Prirm~caschinensis),其掌状深裂的大孢子叶与现代苏铁属十分相似 ,从 而将苏铁起源的时间推到了二叠纪。朱家槽等 (1994)又进一步发现了苏铁小孢子叶球古 生铁花 (Cycadostrobiluspaleozoicus),证明了我国华北地区早在古生代二叠纪就有苏铁的生 长 。最近,胡雨帆等 (Hu等,1999)根据采 自甘肃石炭世苏铁类大孢子叶化石 ,推论苏 铁类植物很可能起源于我国西北地区。管中天 (1996)并认为在古生代华北地区是苏铁类 植物的分布中心 ,但在中生代则移向西南地区。由于西南地区 自古生代以来地质环境和气 候条件一直相对较稳定,可为苏铁植物的进一步发生发展提供良好的条件。此外 ,较为丰 富的苏铁和种子蕨化石的记录 (徐仁等 ,1979)为此提供了古植物学 的证据。从苏铁属的 现代地理分布 卜分析 ,根据王定跃 (Wang,2001)的研究 ,虽然现代苏铁属的二大分布中 心在中国西 南和澳大利亚及其邻近岛屿 ,但我国西南地区至中南半岛地 区是该属原始类群 苏铁亚属的集中分布区域 ,同时也包含了不同进化水平的 3个亚属的类群,因此 ,我国的 西南地 区很可能是现代苏铁的发源地 。 松柏哑门有二纲 ,松柏纲含 6科,我国缺南洋杉科 (Araueariaeeae),贝壳杉科 (Ag— athidaceae),金松 科 (Sciadopityaceae)3科 。尽 管 金松 科 见 于 日本 中部 至南 部 ,不 达 我 国。 但它比中国出现的柏 、杉 、松 3科为古老。 柏科 (Cupressaceae)是从三叠纪到早侏罗纪起源和分化 的两温带分 布大科。在 l8~ 20属中,北温带分布即有 l3属 ,而我 国有其 11,东亚 一北美分布型 2个 ,北温带分布型 3个 ,东亚分布型 4个 。故而科 的分 布型用复合表示 ,应为南 北温带广布 ,即 8—4 (9, 14)型 。其起 源应该 认为 是在 泛 古 大 陆 时 ,古 北 大 陆 东 部 (E.Laurasia),是 逐 渐 向古 南 大陆西部扩散的。我 国该科虽然没有一个特有属 ,却有 2个准特有属 ,1是翠柏属 (Ca/o一 维普资讯 http://www.cqvip.com

维鲁资i讯hitp:/www.cqvip.co5856期吴征镂等:中国植物区系中的特有性及其起源和分化cedrus),从缅、越分布到滇黔桂和云南高原、海南、台湾,台湾或为1变种,因形态和分布星散而显然古老,属7-3型。近来分子系统学证据表明该属同北美西部特有的Heyderiadecurrens(Li,1953)聚在一起(Gadk等,2000),进而归并人翠柏属(Calocedrus)中。因此,该属也视为东亚-北美间断分布。另1属即福建柏属Fokienia,属7-4型,国外仅分布到越南北方、老挝、泰国,是清代还存在的“范范树海”中的主要成员之一,国内则见于福建、浙江、云南等地而且星散,从白垩纪晚期到渐新世以前,在我国新疆、吉林都有化石记录(周浙昆和ArataMomohara,2005),在北美加拿大古新世也有化石出现(Mclver等,1990;Mclver,1992);这些有限的化石记录表明该属有着古老的起源历史,在晚白垩纪到中新世以前曾广布于北半球(周浙昆和ArataMomohara,2005)。杉科(Taxodiaceae)仪是世界有9(-10)属12(-16)种的小科,其所有的属的分布区皆非常狭小,除1属分布于南半球外,余均分布在北半球,均为残遗分布(Li,1978)。中国有4个单、二型属,共5(-6)种,即水杉属(Metasequoia),水松属(Glyptostroleus),杉属(Cunninghamia),秃杉属(台湾杉属)(Tawania)。后两个虽然也可算准特有,秃杉(Taiwaniaflousiana)到邻近的缅甸北部。杉属(Cunninghamia)则越北、老、束均有分布,但后二处实系引种。4属中水杉、水松无疑属古特有,前者且是“水杉区系”(Hu,1980)的表征。从细胞学研究结果看,杉属和秃杉属二属可能系新特有,核型属于IB(李林初,1989)。现代水松(Glyptostrobus)是东亚的第三纪广布的水松种群的残遗种(Florin,1963),通常认为它在杉科中介于北美分布的落羽杉和东亚分布的柳杉间的类群(Liu等,1983;于永福,1995),虽然Gadek等(2000)从形态学并结合分子系统学的研究,认为水松应置于柏科中,建议柏科和杉科合并,从近期的分子系统学的研究也支持这样的的归并(Kusumi等,2000)。但水杉属、水松属,落羽杉属和柳杉属间关系是非常密切的。水杉属在地史上也有着广泛的分布(Chaney,1951),在东亚的残遗无疑均归结于晚第三纪的气候变冷以及第四纪冰期的作用。水杉同北美红杉属(Sequoia),巨杉属(Sequoiadendorn)关系最为密切;它们可能是水松属和北美分布的落羽杉属的祖先类群,水松属同落羽杉属的关系密切,二者在早期的胚胎发育非常相似,同柳杉属构成了一进化关系密切的姐妹群(Kusumi等,2000)。此外,从起源时间上看,杉科化石最早出现于中侏罗世地层,白垩纪几乎所有的类型都出现,并且已广布北半球。但自始新世以后,特别是在晚第三纪和更新世后便相继灭绝(Li等,2003)。杉科的外类群如Voltziaceae的许多类群则出现在晚二叠纪至早侏罗世(Florin,1963),因此,杉科植物起源的时间可追溯到晚三叠世至早侏罗世(于永福,1999)。从化石记录看(Florin,1963),水松属和杉属、水杉属的化石历史非常相似,秃杉的化石较少。杉属有1变种在川西南安宁河谷。而秃杉属有1变种或有2种,1种间断分布于华中和横断山区南段至缅甸北部,另1种极相近而是属模式,孤立于台湾,老树在台湾视为神树。它们的分布格局显然提示台湾从大陆分离可能在喜马拉雅造山运动的后期。本科中除中国特有、准特有4属外,东亚还出现柳杉属(Cryptomeria),含2种,1种在中国,1种在日本。它们如与北美西部的红杉属(Sequoia),巨杉属(Sequoiadendron),北美(东南、西南至中美)的落羽杉属(Taxodium)和孤立于大洋洲塔斯马尼亚的Athrotaxis合在一起看,本科在(三叠纪至)晚白垩纪的分化和分布的轨迹显然是在第一次泛古大陆
6期 吴征镒等 :中国植物区系中的特有性及其起源和分化 585 cedrus),从缅 、越分布到滇黔桂和云南高原 、海南 、台湾 ,台湾或为 1变种 ,因形态和分 布星散而显然古老 ,属 7—3型。近来分子系统学证据表明该属同北美西部特有 的 Heyderia decurrens(Li,1953)聚在一起 (Gadk等 ,2000),进而归并人翠柏属 (Calocedrus)中。因 此 ,该属也视为东亚 一北美间断分布。另 1属即福建柏属 Fokienia,属 7—4型 ,国外仅分 布到越南北方 、老挝 、泰国,是清代还存在的 “茫茫树海 ”中的主要成员之一 ,国内则见 于福建、浙江 、云南等地 而且星散 ,从白垩纪晚期到渐新世以前 ,在我 国新疆 、吉林都有 化石记录 (周浙昆和 ArataMomohara,2005),在北美加拿大古新世也有化石 出现 (Mclver 等,1990;Mclver,1992);这些有限的化石记录表明该属有着古老的起 源历史 ,在晚 白垩 纪到 中新世 以前 曾广 布于北 半球 (周 浙 昆和 ArataMomohara,2005)。 杉科 (Taxodiaceae)虽仪是世界有 9(一10)属 12(一16)种的小科 ,其所有的属的分 布 区皆非常狭小,除 1属分布于南半球外 ,余均分布在北半球 ,均为残遗分 布 (Li,1978)。 中国有 4个单、二型属,共 5(一6)种,即水杉属 (Metasequoia),水松属 (Glyptostroleus),杉属 (Cunninghamia),秃杉属 (台湾杉属)(Taiwania)。后两个虽然也可算准特有,秃杉 (Taiwania /zo )到邻近的缅甸北部。杉属 (Cunninghamia)则越北 、老 、柬均有分布 ,但后二处 实系引种。4属中水杉 、水松无疑属古特有 ,前者且是 “水杉区系” (Hu,1980)的表征。 从 细胞学研 究结 果看 ,杉 属和 秃杉 属二 属可 能系新 特有 ,核 型属 于 liB (李林 初 ,1989)。 现代水松 (Glyptostrobus)是东亚的第 三纪广布 的水松种群 的残 遗种 (Florin.1963), 通常认为它在杉科 中介于北美分 布的落羽杉和东亚分布 的柳杉间的类群 (LJiu等 ,1983; 于永福 ,1995),虽然 Gadek等 (2000)从形态学并结合分子系统学 的研究 ,认 为水松应 置于柏科中,建议 柏科和杉科 合并,从 近期 的分子系统学 的研究也 支持这样 的的归并 (Kusumi等 ,2000)。但水杉属 、水松属 ,落羽杉属和柳杉属问关系是非常密切的。 水杉属住地史上也有着广泛 的分布 (Chaney,1951),在东亚 的残遗无疑均 归结 于晚 第 纪的气候变冷以及第四纪冰期 的作用。水杉 同北美红杉属 (5叼Ⅱo ),巨杉属 (Se— quoiadendom)关系最为密切;它们可能是水松属和北美分布的落羽杉属的祖先类群 ,水松 属同落羽杉属的关系密切 ,二者在早期的胚胎发育非常相似 ,同柳杉属构成了一进化关系 密切的姐妹群 (Kusumi等 ,2000)。此外 ,从起源时问上看,杉科化石最早 出现于中侏 罗 世地层,白垩纪几乎所有的类型都出现 ,并且已广布北半球。但 自始新世以后 ,特别是在 晚第三纪和更新世后便相继 灭绝 (Li等 ,2003)。杉科 的外类群如 Voltziaceae的许 多类群 则出现在晚二叠纪至早侏罗世 (Florin,1963),因此 ,杉科植物起源的时间可追溯到晚三 叠世 至早侏 罗世 (于 永福 ,1999)。 从化石记录看 (Florin,1963),水松属和杉属 、水杉属的化石历史非常相似 ,秃杉 的 化石较少。杉属有 1变种在川西南安宁河谷。而秃杉属有 1变种或有 2种 ,1种间断分布 于华中和横断山区南段至缅甸北部,另 1种极相近而是属模式 ,孤立于台湾 ,老树在台湾 视为神树。它们的分布格局显然提示台湾从大陆分离可能在喜马拉雅造山运动的后期。本 科 中除中国特有 、准特有 4属外 ,东亚还出现柳杉属 (Cryptomer/a),含 2种 ,1种在 中 国 ,1种在 日本 。它 们如 与北 美西 部 的红 杉 属 ( ),巨杉 属 (Seq~oiadendron),北 美 (东南 、西南至中美)的落羽杉属 (Taxodium)和孤立 于大洋洲塔斯马尼亚的 Athrotaxis合 在一起看 ,本科在 (三叠纪至 )晚白垩纪的分化和分布的轨迹显然是在第一次泛古大陆 维普资讯 http://www.cqvip.com

维鲁资int.htp://www.cqvip.cor27卷586云南植物研究上,起源于古北大陆东部(尤其是中国东北和日本),然后扩散至古南大陆东部的情景,其在澳大利亚本土东部绝灭,乃由于该陆块的飘移而旱化后才形成隔离。松科(Pinaceae)是松柏纲中大科,是从晚侏罗纪到白垩纪,即第一次泛古大陆的后期才崛起的,显然较柏科和杉科都年轻。主要分布在北温带。松科植物(包括化石材料)的分布基本上限于北半球,因此,松科植物起源于劳亚古陆是可以肯定的(李楠,1999)。Florin(1963)则推测松科起源于东亚。在世界上12~13属200~220种中,我国有10属95种,属达5/6,而种儿近半数。其中有3个特有属,即金钱松属(Pseudolarix),银杉属(Cathaya)和长苞铁杉属(Nothotsuga),3个属都是单型属。尽管中生代以前,远溯到罗纪至三叠纪,松科可能在当时曾是比较庞大的类群,但中生代后已大部分绝灭,代之而起的是白垩纪至老第三纪起源和分化的近代松科各属。银杉属(Cathaya)的发现,和水杉差不多,也是一个轰动一时的活化石。在地史上,该属在晚白垩纪时,在东亚和北美就有分布,后来在早第三纪时可能是经北大西洋陆桥传播到欧洲,在德国(Eschweiler),东西伯利亚(阿尔丹河谷)等地中新世地层中均有化石报道(Ferguson,1967;Florin,1963)。在新生代时银杉已广布于东亚,欧洲和北美(LiuandBasinger,2000)。晚第三纪气候变冷以及第四纪冰期显然是导致银杉消亡的因素,北美最后的化石记录是在中新世晚期,欧洲是在上新世,中国特有的银杉属植物地史上广布于亚洲种群的残遗(LiuandBasinger,2000)。银杉可能是在落叶松亚科(Laricoideae)中接近黄杉属(Pseudotsuga)(9型分布)的一个大部分已在冰期中绝灭的类型,在我国仅存于川、黔、湘、桂少数残存的原始常绿阔叶林中,分散而孤立,已属濒危。长苞铁杉属(Nothotsuga)是胡先最先分出,后才由C,N.Page发表的,无疑是铁杉属(Tsuga)的近亲,它仅残留分布在南岭的西部的常绿阔叶林中,但可能与北美的Hesperopeuce相对应。金钱松属(Pseudolarix)为中国特有的单型属。零星分布于长江中下游各省温暖地区,西起利川,万县东达浙东,南起湖南衡阳,北至苏南(应俊生,1989),华东、华中许多山区均可见到,且能成小片群落。地史上有广泛的分布。美国中部和西部、欧洲、亚洲中部、我国东北及日本均在第三纪的不同时期发现过该属化石(Farjon,1990;Florin,1963)。根据化石资料推测,该属可能起源于白垩纪中晚期,第三纪时曾发展成为北半球中高纬度广布属(李楠,1999)。在北美和亚洲中部,该属似乎出现后不久便消失了,而在欧洲及日本一直保存到更新世第一次冰期降临(Farjon,1990)。同银杉相似,金钱松也是东亚第三纪种群的残遗分布(LepageandBasinger,1995)。银杉属、金钱松属和长苞铁杉属这3个都显然是占特有属。另外,7-4型3~5种的油杉(Keteleeria),可算准特有。从渐新世到上、中新世广泛分布于北半球,上新世日本也有化石记录,中新世末该属植物在北美消失,欧洲则一直保存到更新世早期(Axelrod1986,1976;Miller,1975;Florin,1963)。在紫杉纲(Taxopsida)中,现存罗汉松目(Podocarpales)和三尖杉目(Cephalotaxales)二目。后者含三尖杉科(Cephalotaxaceae),仅1属三尖杉属9种。如若包括穗花杉属(Amentotaxus)则为2属12种,我国即有8种,故虽为东亚型(14型)分布,而分布中心实在中国本土。三尖杉在东亚-北美的中新世、上新世都有化石,欧洲甚至到中侏罗世。分子系统学研究表明(Cheng等,2000)三尖杉科同红豆杉科为单系,且为姐妹群关系
586 云 南 植 物 研 究 27卷 上 ,起源于古北大陆东部 (尤其是 中国东北和 日本),然后扩散至古 南大陆东部的情景 , 其在澳大利亚本土东部绝火,乃由于该陆块的飘移而旱化后才形成隔离 。 松科 (Pinaceae)是松柏纲中大科 ,是从晚侏罗纪到 白垩纪 ,即第一次泛古大陆的后 期才 崛起 的 ,显 然较 柏科 和杉 科都年 轻 。主 要分 布 在 北 温带 。松 科 植 物 (包 括化 石 材 料 ) 的分布基本上限于北半球 ,因此 ,松科植物起源于劳亚古陆是可以肯定的 (李楠 ,1999)。 F10rin (1963)则 推 测松 科 起源 于 东亚 。在世 界 上 l2~l3属 200~220种 中 ,我 国有 l0属 95种 ,属达 5/6,而种几近半数 。其中有 3个特有属 ,即金钱松属 (Pseudolarix),银杉属 (Cathaya)和长苞铁杉属 (Nothotsuga),3个属都是单型属。尽管 中生代 以前 ,远溯到侏 罗纪至三叠纪 ,松科可能在当时曾是比较庞大的类群 ,但中生代后已大部分绝灭 ,代之而 起的是 白垩纪至老第三纪起源和分化的近代松科各属。 银杉属 (Cathaya)的发现,和水杉差不多 ,也是一个轰动一时的活化石 。在地史上 , 该属在晚白垩纪时,住东亚和北美就有分布,后来在早第三纪时可能是经北大西洋陆桥传 播到欧洲 ,在德国 (Eschweiler),东西伯利亚 (阿尔丹河谷 )等地 中新世地层中均有化石 报道 (Ferguson,1967;Florin,1963)。在新生代 时银杉 已广布于东亚 ,欧洲 和北美 (Lju andBasinger,2000)。晚第 纪气候变冷 以及第 四纪冰期显然是导致银杉消亡 的因素,北 美最后的化石记录是在 中新世晚期,欧洲是在上新世 ,中国特有的银杉属植物地史上广布 于亚洲种群的残遗 (LiuandBasinger,2000)。银杉可能是在落叶松亚科 (Laricoideae)中接 近黄杉属 (Pseudotsuga)(9型分布)的一个大部分 已在冰期中绝灭 的类型 ,在我 国仅存于 川 、黔 、湘 、桂少数残仔的原始常绿阔叶林中,分散而孤立 ,已属濒危 。 长苞铁杉属 (Nothotsuga)是胡先辅最先分 出,后才 由 C.N.Page发表 的,无疑是铁 杉属 (Tsuga)的近亲,它仅残 留分布在南岭 的西部 的常绿 阔叶林 中 ,但 可能与北美 的 Hesperopeuce相对 应 。 金钱松属 (Pseudolarix)为中国特有 的单型属 。零星分布于长江中下游各省温暖地 区, 西起,F,IJJI,万县东达浙东,南起湖南衡阳,北至苏南 (应俊生,1989),华东 、华中许多 山区均可见到,且能成小片群落。地史上有广泛的分布。美国中部和西部 、欧洲、亚洲中 部 、我国东北 及 日本均 在 第 三纪 的不 同时 期 发现 过 该 属 化石 (Faqon,1990;Florin, 1963)。根据化石资料推测 ,该属可能起 源于 白垩纪 中晚期 ,第三纪时曾发展成为北 半球 中高纬度广 属 (李楠,1999)。在北美和亚洲中部 ,该属似乎出现后不久便消失了,而在 欧洲及 日本一直保存到更新世第一次冰期降临 (Farjon,1990)。同银杉相似,金钱松也是东 亚第三纪种群的残遗分布 (LepageandBasinger,1995)。银杉属 、金钱松属和长苞铁杉属这 3 个都显然是古特有属。另外 ,7—4型 3~5种的油杉 (Kete/eer/a),可算准特有。从渐新世 钊上 、中新 f}I=广泛分布于北半球 ,上新世 日本也有化石记录,中新世末该属植物在北美消 失 ,欧洲则一直保存到更新世早期 (Axelrod1986,1976;Miller,1975;FIorin,1963)。 在紫杉纲 (Taxopsida)中,现存罗汉松 目 (Podocarpales)和三尖杉 目 (Cephalotaxales) 二 目。后 者含 三尖杉 科 (Cephalotaxaceae),仅 1属j 尖杉 属 9种。如若 包括 穗 花杉 属 (Amentotaxus)则 为 2属 l2种 ,我 国 即有 8种 ,故 虽 为东业 型 (14型 )分 布 ,而分 布 中 心 实在中国本土。三尖杉在东亚 一北美的中新世 、上新世都有化石 ,欧洲甚至到中侏罗世。 分子系统学研究表明 (Cheng等 ,2000)三尖杉科同红豆杉科为单系 ,且为姐妹群关系, 维普资讯 http://www.cqvip.com