
二、细胞壁以内的构造—原生质体 l、细胞质膜(cytoplasmic membrane) 又称质膜(plasma membrane))、细胞膜(cell membrane)或内膜(inner membrane),是紧贴在细胞壁内 侧、包围着细胞质的一层柔软、脆弱、富有弹性的半透性薄膜,厚约7~8m,由碳脂(占20%~30%)和 蛋白质(占50%一70%)组成。通过质壁分离、鉴别性染色或原生质体破裂等方法可在光学显微镜下观察到: 用电子显微镜观察细菌的超薄切片,则可更清楚地观察到它的存在。电镜观察到的细胞质膜,是在上下两 暗色层之间夹着一浅色中间层的双层膜结构。这是因为,组成细胞膜主要成分的磷脂,是由两层磷脂分子 按一定规律整齐地排列而成的。其中每一个磷胎分子由一个带正电荷日能溶于水的极性头(磷酸端)和 个不带电荷、不溶于水的非极性尾(烃端)所构成。极性头朝向内外两表面,呈亲水性,而非极性端的疏 水尾则埋入膜的内层,于是形成了一个磷脂双分子层。在极性头的甘油3C上,不同种微生物其有不同的R 基,如磷脂酸、磷脂酰甘油、磷脂酰乙醇胺、磷脂酰胆碱、磷脂酰丝氨酸或磷脂酰肌醇等(图3一10)。在 原核微生物的细胞质膜上多数含磷脂酰甘油,此外,在革兰氏阴性细菌中,多数还含磷脂酰乙醇胺,在分 枝杆菌中则含磷脂酰肌醇,等等。而非极性尾则由长链脂肪酸通过酯键连接在甘油的C1和2位上组成, 其链长和饱和度因细菌种类和生长温度而异,通常生长温度要求越高的种,其饱和度也越高,反之则低, 有关细胞质膜的结构与功能的解释,较多的学者仍倾向于1972年由辛格(J.S.Singer)和尼科尔森 (G.L.Nicolson)所提出的液态镶嵌模型(fluid mosaic model)。其要点为:①膜的主体是脂质双分子层: ②脂质双分子层具有流动性:③整合蛋白因其表面呈疏水性,故可“溶”于脂质双分子层的疏水性内层中: ④周边蛋白表面含有亲水基团,故可通过静电引力与脂质双分子层表面的极性头相连:⑤脂质分子间或脂 质与蛋白质分子间无共价结合:©脂质双分子层犹如一“海洋”,周边蛋白可在其上作“漂浮”运动,而 整合蛋白则似“冰山”状沉浸在其中作横向移动。有关细胞质膜的模式构造可见图3一1。 细胞膜的生理功能为: ①选择性地控制细胞内、外的营养物质和代谢产物的运送 ②是维持细胞内正常泰透压的屏障: ③合成细胞壁和糖被的各种组分(肽聚糖、磷壁酸、LPS、荚膜多糖等)的重要基地 ④膜上含有氧化磷酸化或光合磷酸化等能量代谢的酶系,是细胞的产能场所: ⑤是鞭毛基体的着生部位和鞭毛旋转的供能部位。 原核微生物的细胞质膜上一般不含胆固醇等甾醇,这一点与真核生物明显不同。但缺乏细胞壁的原核 生物 -枝原体(Mycoplasma)则属例外。在其细胞膜上因含有hopanoid类甾醇而增强了坚韧性,故在 一定程度上弥补了因缺壁而带来的不足。多烯类抗生素因可破坏含甾醇的细胞质膜,故可抑制枝原体和真 核生物,但对其他的原核生物则无抑制作用。 间体(msos0me,或中体)是一种由细胞质膜内褶而形成的囊状构造,其中充满着层状或管状的泡囊。 多见于革兰氏阳性细菌。每个细胞含一至少数几个。着生部位可在表层或深层,前者与某些醇如青霉素酶 的分泌有关,后者与DNA的复制、分配以及与细胞分裂有关。近年来也有学者提出不同的看法,认为“间 体”仅是电镜制片时因脱水操作而引起的一种惯像。 2、细胞质和内含物 细胞质(cytoplasm)是细胞质膜包围的除核区外的一切半透明、胶状、颗粒状物质的总称。含水量 约80%。原核微生物的细胞质是不流动的,这一点与真核生物明显不同。细胞质的主要成分为核糖体(由 50S大亚基和30S小亚基组成)、贮藏物、多种酶类和中向代谢物、质粒、各种营养物和大分子的单体等
25 二、细胞壁以内的构造——原生质体 1、细胞质膜(cytoplasmic membrane) 又称质膜(plasma membrane)、细胞膜(cell membrane)或内膜(inner membrane),是紧贴在细胞壁内 侧、包围着细胞质的一层柔软、脆弱、富有弹性的半透性薄膜,厚约 7~8nm,由磷脂(占 20%~30%)和 蛋白质(占 50%~70%)组成。通过质壁分离、鉴别性染色或原生质体破裂等方法可在光学显微镜下观察到; 用电子显微镜观察细菌的超薄切片,则可更清楚地观察到它的存在。电镜观察到的细胞质膜,是在上下两 暗色层之间夹着一浅色中间层的双层膜结构。这是因为,组成细胞膜主要成分的磷脂,是由两层磷脂分子 按一定规律整齐地排列而成的。其中每一个磷脂分子由一个带正电荷且能溶于水的极性头(磷酸端)和一 个不带电荷、不溶于水的非极性尾(烃端)所构成。极性头朝向内外两表面,呈亲水性,而非极性端的疏 水尾则埋入膜的内层,于是形成了一个磷脂双分子层。在极性头的甘油 3C 上,不同种微生物具有不同的 R 基,如磷脂酸、磷脂酰甘油、磷脂酰乙醇胺、磷脂酰胆碱、磷脂酰丝氨酸或磷脂酰肌醇等(图 3-10)。在 原核微生物的细胞质膜上多数含磷脂酰甘油,此外,在革兰氏阴性细菌中,多数还含磷脂酰乙醇胺,在分 枝杆菌中则含磷脂酰肌醇,等等。而非极性尾则由长链脂肪酸通过酯键连接在甘油的 C1 和 C2 位上组成, 其链长和饱和度因细菌种类和生长温度而异,通常生长温度要求越高的种,其饱和度也越高,反之则低。 有关细胞质膜的结构与功能的解释,较多的学者仍倾向于 1972 年由辛格(J.S.Singer)和尼科尔森 (G.L.Nicolson)所提出的液态镶嵌模型(fluid mosaic model)。其要点为:①膜的主体是脂质双分子层; ②脂质双分子层具有流动性;③整合蛋白因其表面呈疏水性,故可“溶”于脂质双分子层的疏水性内层中; ④周边蛋白表面含有亲水基团,故可通过静电引力与脂质双分子层表面的极性头相连;⑤脂质分子间或脂 质与蛋白质分子间无共价结合;⑥脂质双分子层犹如一“海洋”,周边蛋白可在其上作“漂浮”运动,而 整合蛋白则似“冰山”状沉浸在其中作横向移动。有关细胞质膜的模式构造可见图 3-11。 细胞膜的生理功能为: ①选择性地控制细胞内、外的营养物质和代谢产物的运送; ②是维持细胞内正常渗透压的屏障; ③合成细胞壁和糖被的各种组分(肽聚糖、磷壁酸、LPS、荚膜多糖等)的重要基地; ④膜上含有氧化磷酸化或光合磷酸化等能量代谢的酶系,是细胞的产能场所; ⑤是鞭毛基体的着生部位和鞭毛旋转的供能部位。 原核微生物的细胞质膜上一般不含胆固醇等甾醇,这一点与真核生物明显不同。但缺乏细胞壁的原核 生物——枝原体(Mycoplasma)则属例外。在其细胞膜上因含有 hopanoid 类甾醇而增强了坚韧性,故在 一定程度上弥补了因缺壁而带来的不足。多烯类抗生素因可破坏含甾醇的细胞质膜,故可抑制枝原体和真 核生物,但对其他的原核生物则无抑制作用。 间体(mesosome,或中体)是一种由细胞质膜内褶而形成的囊状构造,其中充满着层状或管状的泡囊。 多见于革兰氏阳性细菌。每个细胞含一至少数几个。着生部位可在表层或深层,前者与某些酶如青霉素酶 的分泌有关,后者与 DNA 的复制、分配以及与细胞分裂有关。近年来也有学者提出不同的看法,认为“间 体”仅是电镜制片时因脱水操作而引起的一种赝像。 2、细胞质和内含物 细胞质(cytoplasm)是细胞质膜包围的除核区外的一切半透明、胶状、颗粒状物质的总称。含水量 约 80%。原核微生物的细胞质是不流动的,这一点与真核生物明显不同。细胞质的主要成分为核糖体(由 50S 大亚基和 30S 小亚基组成)、贮藏物、多种酶类和中间代谢物、质粒、各种营养物和大分子的单体等

少数细菌还有类囊体、羧酶体、气泡或伴孢晶体等。细胞质内形状较大的颗粒状构造称为内含物(inclusion body),包括各种贮藏物和羧酶体、气泡等。 (I)贮藏物(reserve materials) 贮藏物是一类由不同化学成分累积而成的不溶性沉淀颗粒,主要功能是贮存营养物。种类很多,表解 如下: 糖原:大肠杆菌、克雷伯氏菌、芽孢杆菌和蓝细菌等 碳源及能源类聚B-羟丁酸(PHB):固氨菌、产碱菌和肠杆菌等 硫粒:紫硫细菌、丝硫细菌、贝氏硫杆菌等 贮藏物 氨源类 藻青素:蓝细菌 藻青蛋白:蓝细菌 磷源(异染粒):迁回螺菌、白喉棒杆菌、结核分枝杆菌 ①聚-B-轻丁酸(poly-B-hydroxybutyrate,.PHB):是存在于许多细菌细胞质内属于类脂性质的碳 源类贮藏物,不溶于水,可溶于氯仿,可用尼罗蓝或苏丹黑染色,具有贮藏能量、碳源和降低细胞内渗透 压的作用。当巨大芽孢杆菌在含乙酸或丁酸的培养基中生长时,细胞内贮藏的PHB可达其干重的6O%。在 棕色周氨菌的孢囊中也含PHB。 PB于1929年被发现,至今已发现60属以上的细菌能合成并贮藏。由于它无毒、可塑、易降解 故认为是生产医用塑料、生物降解塑料的良好原料。若干产碱菌、固氮菌和假单胞菌是主要的生产菌 种。近年来,又发现在一些革兰氏阳性和阴性好氧菌、光合厌氧细菌中,都存在PHB类化合物,它们与PH 仅是R基不同(R=CH时即为HB)。这类化合物可统称为聚羟链烷酸 ②多糖类贮藏物:包括糖原和淀粉类。在真细菌中以糖原为多。糖原可用碘液染成褐色,在光学显微 镜下可见。 ③异染粒(■etachromatic granules):义称迂回体或捩转菌素(volutin granules),这是因为它最早 是在迂回螺菌被发现并可用美蓝或甲苯胺蓝染成红紫色的缘故。颗粒大小为0.5~1.0μm,是无机偏磷酸 的聚合物,分子呈线状,n值在2一10间。一般在含磷丰富的环境下形成。功能是贮藏磷元素和能量,并 可降低细胞的渗透压。在白喉棒杆菌和结核分枝杆菌中极易见到,因此可用于有关细菌的鉴定。异染粒 的化学结构为: ④藻青素(cya ophycin):通常存在于蓝细菌中,是一种内源性源贮藏物,同时还兼有贮存 能源的作用。一般呈颗粒状,由含精氨酸和天冬氨酸残基(1:1)的分枝多肽所构成,分子量在25,000~ 125,000范围内。 (2)磁小体(egnetosome)197 勃莱克摩(R.P.Blakemore)在一种称为折叠螺旋体的趋磁细南中发现。目前所知的趋磁细菌主要为 水生螺菌属和嗜胆球菌属中。这些细菌细胞中含有大小均匀、数目不等的磁小体,其成分为下阳0:,外有 一层磷脂、蛋白或糖蛋白膜包裹,是单磁砖晶体,无毒,大小均匀(20~100m),每个细胞内有2~20 颗。形状为平截八面体、平行六面体或六棱柱体等。其功能是导向作用,即借鞭毛游向对该菌最有利的泥、 水界面微氧环境处生活。目前认为趋磁菌有一定的实用前景,包括生产磁性定向药物或抗体,以及制造生 物传感器等。 (3)羧酶体(carboxysome 36
26 少数细菌还有类囊体、羧酶体、气泡或伴孢晶体等。细胞质内形状较大的颗粒状构造称为内含物(inclusion body),包括各种贮藏物和羧酶体、气泡等。 (1)贮藏物(reserve materials) 贮藏物是一类由不同化学成分累积而成的不溶性沉淀颗粒,主要功能是贮存营养物。种类很多,表解 如下: ①聚-β-羟丁酸(poly-β-hydroxybutyrate, PHB):是存在于许多细菌细胞质内属于类脂性质的碳 源类贮藏物,不溶于水,可溶于氯仿,可用尼罗蓝或苏丹黑染色,具有贮藏能量、碳源和降低细胞内渗透 压的作用。当巨大芽孢杆菌 在含乙酸或丁酸的培养基中生长时,细胞内贮藏的 PHB 可达其干重的 60%。在 棕色固氮菌 的孢囊中也含 PHB。 PHB 于 1929 年被发现,至今已发现 60 属以上的细菌能合成并贮藏。由于它无毒、可塑、易降解, 故认为是生产医用塑料、生物降解塑料的良好原料。若干产碱菌 、固氮菌 和假单胞菌 是主要的生产菌 种。近年来,又发现在一些革兰氏阳性和阴性好氧菌、光合厌氧细菌中,都存在 PHB 类化合物,它们与 PHB 仅是 R 基不同(R=CH3 时即为 PHB)。这类化合物可统称为聚羟链烷酸 。 ②多糖类贮藏物:包括糖原和淀粉类。在真细菌中以糖原为多。糖原可用碘液染成褐色,在光学显微 镜下可见。 ③异染粒(metachromatic granules):又称迂回体或捩转菌素(volutin granules),这是因为它最早 是在迂回螺菌 被发现并可用美蓝或甲苯胺蓝染成红紫色的缘故。颗粒大小为 0.5~1.0μm,是无机偏磷酸 的聚合物,分子呈线状,n 值在 2~106 间。一般在含磷丰富的环境下形成。功能是贮藏磷元素和能量,并 可降低细胞的渗透压。在白喉棒杆菌 和结核分枝杆菌 中极易见到,因此可用于有关细菌的鉴定。异染粒 的化学结构为: ④藻青素(cyanophycin):通常存在于蓝细菌中,是一种内源性氮源贮藏物,同时还兼有贮存 能源的作用。一般呈颗粒状,由含精氨酸和天冬氨酸残基(1:1)的分枝多肽所构成,分子量在 25,000~ 125,000 范围内。 (2)磁小体(megnetosome) 197 勃莱克摩(R.P.Blakemore)在一种称为折叠螺旋体 的趋磁细菌中发现。目前所知的趋磁细菌主要为 水生螺菌属 和嗜胆球菌属 中。这些细菌细胞中含有大小均匀、数目不等的磁小体,其成分为 Fe3O4,外有 一层磷脂、蛋白或糖蛋白膜包裹,是单磁畴晶体,无毒,大小均匀(20~100nm),每个细胞内有 2~20 颗。形状为平截八面体、平行六面体或六棱柱体等。其功能是导向作用,即借鞭毛游向对该菌最有利的泥、 水界面微氧环境处生活。目前认为趋磁菌有一定的实用前景,包括生产磁性定向药物或抗体,以及制造生 物传感器等。 (3)羧酶体(carboxysome)

又称羧化体,是存在于一些自养细菌细胞内的多角形或六角形内含物。其大小与噬菌体相仿,约10m, 内含1,5-二磷酸核酮糖羧化酶,在自养细菌的00,固定中起者关键作用。在排硫硫杆南、那不勒斯破杆菌 贝日阿托氏菌属、硝化细菌和一些蓝细茵中均可找到羧酶体。 (4)湘(gasv0cu01es) 是在许多光合营养型、无鞭毛运动的水生细菌中存在的充满气体的泡囊状内含物,大小为0.2一1.0 μm×75m,内由数排柱形小空泡组成,外有2nm厚的蛋白质膜包裹,其功能是调节细胞比重以使细胞漂 浮在最适水层中获取光能、0,和营养物质。每个细胞含几个至几百个气泡。如鱼腥蓝细菌属、顶孢蓝细菌 属、盐杆菌属、暗网菌属和红假单胞菌的一些种中都有气泡。 3、核区(nuclear region or area) 又称核质体(nuclear body)、原核、拟核(nucleoid)或核基因组(genome)。指原核生物所特有的无 核膜结构、无固定形态的原始细胞核。用富尔根(Feulger)染色法染色后,可见到呈紫色的形态不定的核 区。它是一个大型环状双链NA分子,只有少量蛋白质与之结合,长度一般为0.25~3.00m,例如,大肠 杆菌的核区DNA长约1.1~1.4mm,枯草芽孢杆菌的约为1.7mm,嗜血流感杆菌约0.832mm。每个细胞所含 的核区数与该细菌的生长速度有关, 一般为1~4个。在快速生长的细菌中,核区DNA可占细胞总体积的 20%。细菌的核区除在染色体复制的短时间内呈双倍体外,一般均为单倍体。核区是细菌负载遗传信息的 主要物质基础 4、特殊的休眠构造一芽孢 某些细菌在其生长发育后期,在细胞内形成一个圆形或椭圆形、厚壁、含水量极低、抗逆性极强的休 眠体,称为芽孢(endospore或spore,.偶译“内生孢子”)。每一营养细胞内仅生成一个芽孢。芽孢是 整个生物界中抗逆性最强的生命体,在抗热、抗化学药物、抗辐射和抗静水压等方面,更是首屈一指。 般细菌的营养细胞不能经受70℃以上的高温,可是,它们的芽孢却有惊人的耐高温能力。例如,肉毒梭菌 的芽孢在100℃沸水中要经过5.0一9.5h才被杀死,至121℃时,平均也要10min才杀死:热解糖梭菌的 营养细胞在50℃下经数分钟即可杀死,但它的一群芽孢却须在132℃下经4.4mi才能杀死其中的90%。芽 孢的抗紫外线能力一般是其营养细胞的一倍。巨大芽孢杆菌芽孢的抗辐射能力要比大肠杆菌的营养细胞强 36倍。芽孢的休眠能力更是突出。在其休眠期间,不能检查出任何代谢活力,因此称为隐生态 (cryptobiosis)。 一般的芽孢在普通的条件下可保持几年至几十年的生活力。但文献中还有许多更突出 的记载,如:环状芽孢杆菌的芽孢在植物标本上(英国)己保存200~300年: 一种高温放线菌的芽孢 在建筑材料中(美国)已保存2000年:普通高温放线菌的芽孢在湖底冻土中(美国)已保存7500年 一种芽孢杆南(aci11ussD)的芽孢在琥珀内蜜蜂肠道中(美国)已保存2500万-4000万年。 (1)产芽孢细菌的种类 能产芽孢的细菌属不多,最主要的是属于革兰氏阳性杆菌的两个属一一好氧性的芽孢杆菌属和厌氧 性的梭菌属。球菌中只有芽孢八叠球菌属产生芽孢。螺菌中的孢螺菌属也产芽孢。此外,还发现少数 其他杆南可产生芽孢,如芽孢乳杆菌属、脱硫肠状菌属、考克斯氏体属(Coxiella)、鼠孢南属和高温 放线菌属等。芽孢的有无、形态、大小和着生位置是细菌分类和鉴定中的重要指标。 (2)芽和的构浩 从图3-13和以下的表解中就可以较清楚地了解芽孢的细致构造和主要功能
27 又称羧化体,是存在于一些自养细菌细胞内的多角形或六角形内含物。其大小与噬菌体相仿,约 10nm, 内含 1,5-二磷酸核酮糖羧化酶,在自养细菌的 CO2 固定中起着关键作用。在排硫硫杆菌 、那不勒斯硫杆菌 、 贝日阿托氏菌属 、硝化细菌和一些蓝细菌中均可找到羧酶体。 (4)气泡(gas vocuoles) 是在许多光合营养型、无鞭毛运动的水生细菌中存在的充满气体的泡囊状内含物,大小为 0.2~1.0 μm×75nm,内由数排柱形小空泡组成,外有 2nm 厚的蛋白质膜包裹,其功能是调节细胞比重以使细胞漂 浮在最适水层中获取光能、O2 和营养物质。每个细胞含几个至几百个气泡。如鱼腥蓝细菌属 、顶孢蓝细菌 属 、盐杆菌属 、暗网菌属 和红假单胞菌 的一些种中都有气泡。 3、核区(nuclear region or area) 又称核质体(nuclear body)、原核 、拟核(nucleoid)或核基因组(genome)。指原核生物所特有的无 核膜结构、无固定形态的原始细胞核。用富尔根(Feulgen)染色法染色后,可见到呈紫色的形态不定的核 区。它是一个大型环状双链 DNA 分子,只有少量蛋白质与之结合,长度一般为 0.25~3.00mm,例如,大肠 杆菌的核区 DNA 长约 1.1~1.4mm,枯草芽孢杆菌的约为 1.7mm,嗜血流感杆菌 约 0.832mm。每个细胞所含 的核区数与该细菌的生长速度有关,一般为 1~4 个。在快速生长的细菌中,核区 DNA 可占细胞总体积的 20%。细菌的核区除在染色体复制的短时间内呈双倍体外,一般均为单倍体。核区是细菌负载遗传信息的 主要物质基础。 4、特殊的休眠构造——芽孢 某些细菌在其生长发育后期,在细胞内形成一个圆形或椭圆形、厚壁、含水量极低、抗逆性极强的休 眠体,称为芽孢(endospore 或 spore,偶译“内生孢子”)。每一营养细胞内仅生成一个芽孢。芽孢是 整个生物界中抗逆性最强的生命体,在抗热、抗化学药物、抗辐射和抗静水压等方面,更是首屈一指。一 般细菌的营养细胞不能经受 70℃以上的高温,可是,它们的芽孢却有惊人的耐高温能力。例如,肉毒梭菌 的芽孢在 100℃沸水中要经过 5.0~9.5h 才被杀死,至 121℃时,平均也要 10min 才杀死:热解糖梭菌 的 营养细胞在 50℃下经数分钟即可杀死,但它的一群芽孢却须在 132℃下经 4.4min 才能杀死其中的 90%。芽 孢的抗紫外线能力一般是其营养细胞的一倍。巨大芽孢杆菌芽孢的抗辐射能力要比大肠杆菌的营养细胞强 36 倍。芽孢的休眠能力更是突出。在其休眠期间,不能检查出任何代谢活力,因此称为隐生态 (cryptobiosis)。一般的芽孢在普通的条件下可保持几年至几十年的生活力。但文献中还有许多更突出 的记载,如:环状芽孢杆菌 的芽孢在植物标本上(英国)已保存 200~300 年;一种高温放线菌 的芽孢 在建筑材料中(美国)已保存 2000 年;普通高温放线菌 的芽孢在湖底冻土中(美国)已保存 7500 年; 一种芽孢杆菌(Bacillus sp) 的芽孢在琥珀内蜜蜂肠道中(美国)已保存 2500 万~4000 万年。 (1)产芽孢细菌的种类 能产芽孢的细菌属不多,最主要的是属于革兰氏阳性杆菌的两个属——好氧性的芽孢杆菌属 和厌氧 性的梭菌属 。球菌中只有芽孢八叠球菌属 产生芽孢。螺菌中的孢螺菌属 也产芽孢。此外,还发现少数 其他杆菌可产生芽孢,如芽孢乳杆菌属 、脱硫肠状菌属 、考克斯氏体属(Coxiella)、鼠孢菌属 和高温 放线菌属 等。芽孢的有无、形态、大小和着生位置是细菌分类和鉴定中的重要指标。 (2)芽孢的构造 从图 3-13 和以下的表解中就可以较清楚地了解芽孢的细致构造和主要功能

芽孢衣 皮层 芽孢壁 芽孢 核心 芽孢质 一芽孢核区 一孢外壁 图3-13细菌芽孢构造模式图 图3-13细菌芽孢构造模式图 芽孢囊:是产芽孢蘭的营养细胞外壳 产芽孢细菌 (孢外壁:主要含脂蛋白,透性差(有的芽孢无此层) 芽孢衣:主要含硫水性角蛋白抗酶解、抗药物,多价阳离子难通过 芽孢皮层:主要含芽孢肽聚糖及DPA-Ca,体积大,渗透压高 芽孢壁:含肽聚糖,可发展成新细胞的壁 核心车孢质膜:含磷脂、蛋白质,可发展成新细胞的膜 芽孢质:含DPA-Ca、核糖体,RNA和酶类 核区:含DNA 皮层(cortex)在芽孢中占有很大体积(36%~60%),内含大量为芽孢皮层所特有的芽孢肚聚糖,其特 点是呈纤维束状、交联度小、负电荷强、可被溶菌酶水解。此外,皮层中还含有占芽孢干重?%一10%的吡 啶二羧酸钙盐(calcium picolinate,DP-Ca),但不含磷壁酸。皮层的渗透压可高达20个大气压左右, 含水量约70%,略低于营养细胞(约80%),但比芽孢整体的平均含水量(40%左右)高出许多。芽孢的核 心(cor©)又称芽孢原生质体,由芽孢壁、芽孢质膜、芽孢质和核区四部分组成,它的含水量极低(10%一 25%),因而特别有利于抗热、抗化学药物(如H0),并可避免酶的失活。除芽孢壁中不含磷壁酸以及芽 孢质中含DPA-Ca外,核心中的其他成分与一般细胞相似。图3-14示芽孢特有的芽孢肚聚糖和吡啶-2,6- 二羧酸钙盐的分子构造。 产芽孢的细菌当其细胞停止生长即环境中缺乏营养及有害代谢产物积累过多时,就开始形成芽孢 从形态上来看,芽孢形成可分七个阶段(图3-15): ①NA浓缩,束状染色质形成 ②细胞膜内陷,细胞发生不对称分裂,其中小体积部分即为前芽孢(forespore): ③前芽孢的双层隔膜形成,这时芽孢的抗辐射性提高: ④在上述两层隔膜间充填芽孢肽聚糖后,合成DPA,紫积钙离子,开始形成皮层,再经脱水,使折光 率增高: ⑤芽孢农合成结束: ⑥皮层合成完成,芽孢成熟,抗热性出现: 28
28 图 3-13 细菌芽孢构造模式图 皮层(cortex)在芽孢中占有很大体积(36%~60%),内含大量为芽孢皮层所特有的芽孢肽聚糖,其特 点是呈纤维束状、交联度小、负电荷强、可被溶菌酶水解。此外,皮层中还含有占芽孢干重 7%~10%的吡 啶二羧酸钙盐(calcium picolinate, DPA-Ca),但不含磷壁酸。皮层的渗透压可高达 20 个大气压左右, 含水量约 70%,略低于营养细胞(约 80%),但比芽孢整体的平均含水量(40%左右)高出许多。芽孢的核 心(core)又称芽孢原生质体,由芽孢壁、芽孢质膜、芽孢质和核区四部分组成,它的含水量极低(10%~ 25%),因而特别有利于抗热、抗化学药物(如 H2O2),并可避免酶的失活。除芽孢壁中不含磷壁酸以及芽 孢质中含 DPA-Ca 外,核心中的其他成分与一般细胞相似。图 3-14 示芽孢特有的芽孢肽聚糖和吡啶-2,6- 二羧酸钙盐的分子构造。 产芽孢的细菌当其细胞停止生长即环境中缺乏营养及有害代谢产物积累过多时,就开始形成芽孢。 从形态上来看,芽孢形成可分七个阶段(图 3-15): ①DNA 浓缩,束状染色质形成; ②细胞膜内陷,细胞发生不对称分裂,其中小体积部分即为前芽孢(forespore); ③前芽孢的双层隔膜形成,这时芽孢的抗辐射性提高; ④在上述两层隔膜间充填芽孢肽聚糖后,合成 DPA,累积钙离子,开始形成皮层,再经脱水,使折光 率增高; ⑤芽孢衣合成结束; ⑥皮层合成完成,芽孢成熟,抗热性出现;
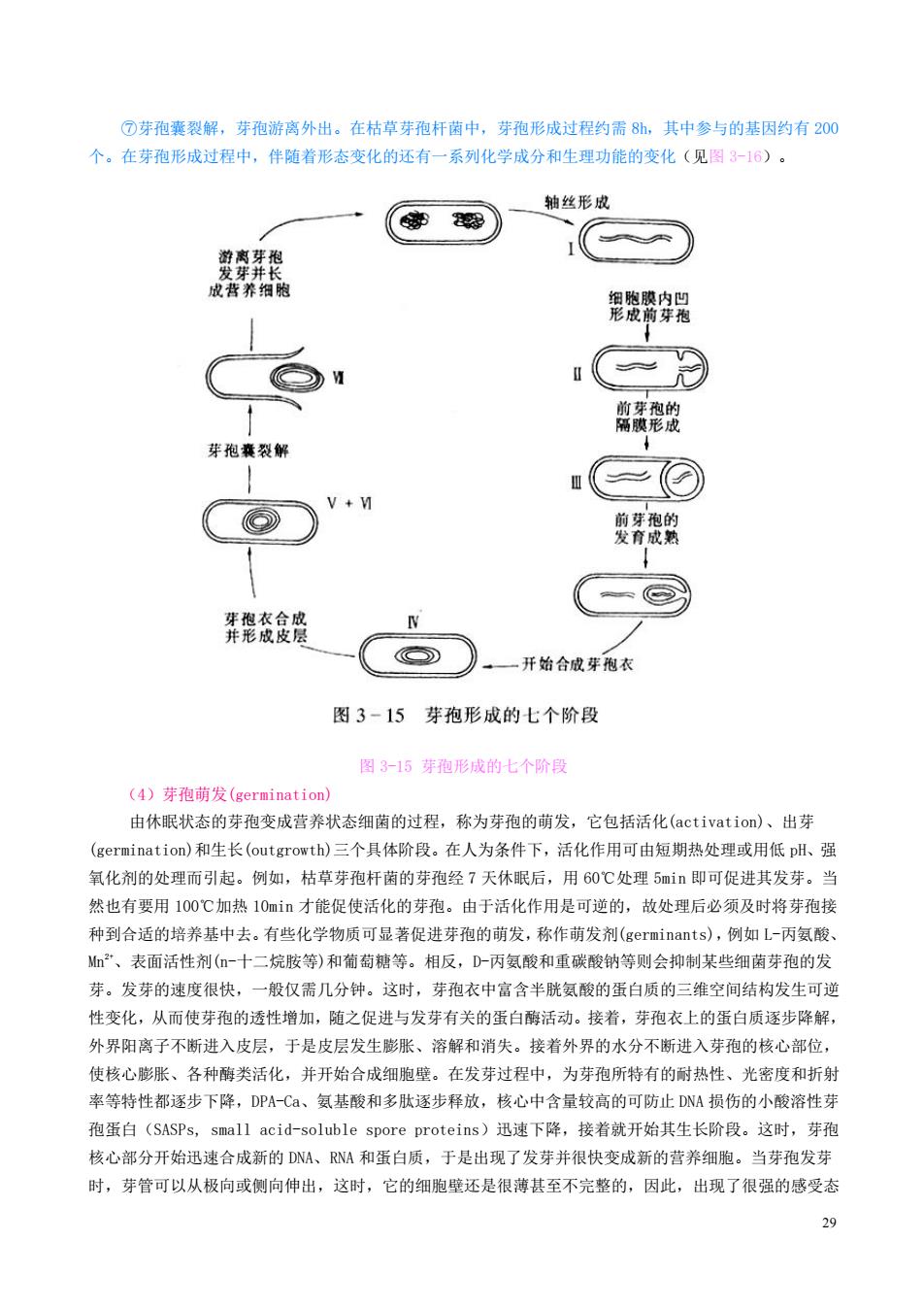
⑦芽孢囊裂解,芽孢游离外出。在枯草芽孢杆菌中,芽孢形成过程约需8h,其中参与的基因约有20 个。在芽孢形成过程中,伴随着形态变化的还有一系列化学成分和生理功能的变化(见图3-16)。 轴丝形成 德器) 郑险穿器 ○M 邮黄形的 芽孢囊裂解 O 牙孢的 弄凳衣食贷 W 开始合成芽孢衣 图3-15芽孢形成的七个阶段 图315芽孢形成的七个阶段 (4)芽孢萌发(germination) 由休眠状态的芽孢变成营养状态细菌的过程,称为芽孢的萌发,它包括活化(activation))、出芽 (germination))和生长(outgrowth)三个具体阶段。在人为条件下,活化作用可由短期热处理或用低plH、强 氧化剂的处理而引起。例如,枯草芽孢杆南的芽孢经7天休眠后,用60℃处理5mi即可促进其发芽。当 然也有要用100℃加热10mi才能促使活化的芽孢。由于活化作用是可逆的,故处理后必须及时将芽孢接 种到合话的培养基中去。有些化学物质可显著促进芽孢的萌发,称作萌发剂(germinants),例如L-丙氨酸 如”、表面活性剂(一十二烷胺等)和葡萄糖等。相反,D一丙氨酸和重碳酸钠等则会抑制某些细菌芽孢的发 芽。发芽的速度很快,一般仅需几分钟。这时,芽孢衣中富含半胱氨酸的蛋白质的三维空间结构发生可逆 性变化,从而使芽孢的透性增加,随之促进与发芽有关的蛋白酶活动。接着,芽孢衣上的蛋白质逐步降解。 外界阳离子不断讲入皮层,于是皮层发生膨张、溶解和消失。接若外界的水分不断讲入芽孢的核心部位」 使核心膨胀、各种酶类活化,并开始合成细胞壁。在发芽过程中,为芽孢所特有的耐热性、光密度和折射 率等特性都逐步下降,DPA-Ca、氨基酸和多肽逐步释放,核心中含量较高的可防止DNA损伤的小酸溶性芽 孢蛋白(SASPs,small acid-soluble spore proteins)迅速下降,接着就开始其生长阶段。这时,芽孢 核心部分开始迅速合成新的DA、A和蛋白质,于是出现了发芽并很快变成新的营养细胞。当芽孢发芽 时,芽管可以从极向或侧向伸出,这时,它的细胞壁还是很薄甚至不完整的,因此,出现了很强的感受态
29 ⑦芽孢囊裂解,芽孢游离外出。在枯草芽孢杆菌中,芽孢形成过程约需 8h,其中参与的基因约有 200 个。在芽孢形成过程中,伴随着形态变化的还有一系列化学成分和生理功能的变化(见图 3-16)。 图 3-15 芽孢形成的七个阶段 (4)芽孢萌发(germination) 由休眠状态的芽孢变成营养状态细菌的过程,称为芽孢的萌发,它包括活化(activation)、出芽 (germination)和生长(outgrowth)三个具体阶段。在人为条件下,活化作用可由短期热处理或用低 pH、强 氧化剂的处理而引起。例如,枯草芽孢杆菌的芽孢经 7 天休眠后,用 60℃处理 5min 即可促进其发芽。当 然也有要用 100℃加热 10min 才能促使活化的芽孢。由于活化作用是可逆的,故处理后必须及时将芽孢接 种到合适的培养基中去。有些化学物质可显著促进芽孢的萌发,称作萌发剂(germinants),例如 L-丙氨酸、 Mn2+、表面活性剂(n-十二烷胺等)和葡萄糖等。相反,D-丙氨酸和重碳酸钠等则会抑制某些细菌芽孢的发 芽。发芽的速度很快,一般仅需几分钟。这时,芽孢衣中富含半胱氨酸的蛋白质的三维空间结构发生可逆 性变化,从而使芽孢的透性增加,随之促进与发芽有关的蛋白酶活动。接着,芽孢衣上的蛋白质逐步降解, 外界阳离子不断进入皮层,于是皮层发生膨胀、溶解和消失。接着外界的水分不断进入芽孢的核心部位, 使核心膨胀、各种酶类活化,并开始合成细胞壁。在发芽过程中,为芽孢所特有的耐热性、光密度和折射 率等特性都逐步下降,DPA-Ca、氨基酸和多肽逐步释放,核心中含量较高的可防止 DNA 损伤的小酸溶性芽 孢蛋白(SASPs, small acid-soluble spore proteins)迅速下降,接着就开始其生长阶段。这时,芽孢 核心部分开始迅速合成新的 DNA、RNA 和蛋白质,于是出现了发芽并很快变成新的营养细胞。当芽孢发芽 时,芽管可以从极向或侧向伸出,这时,它的细胞壁还是很薄甚至不完整的,因此,出现了很强的感受态