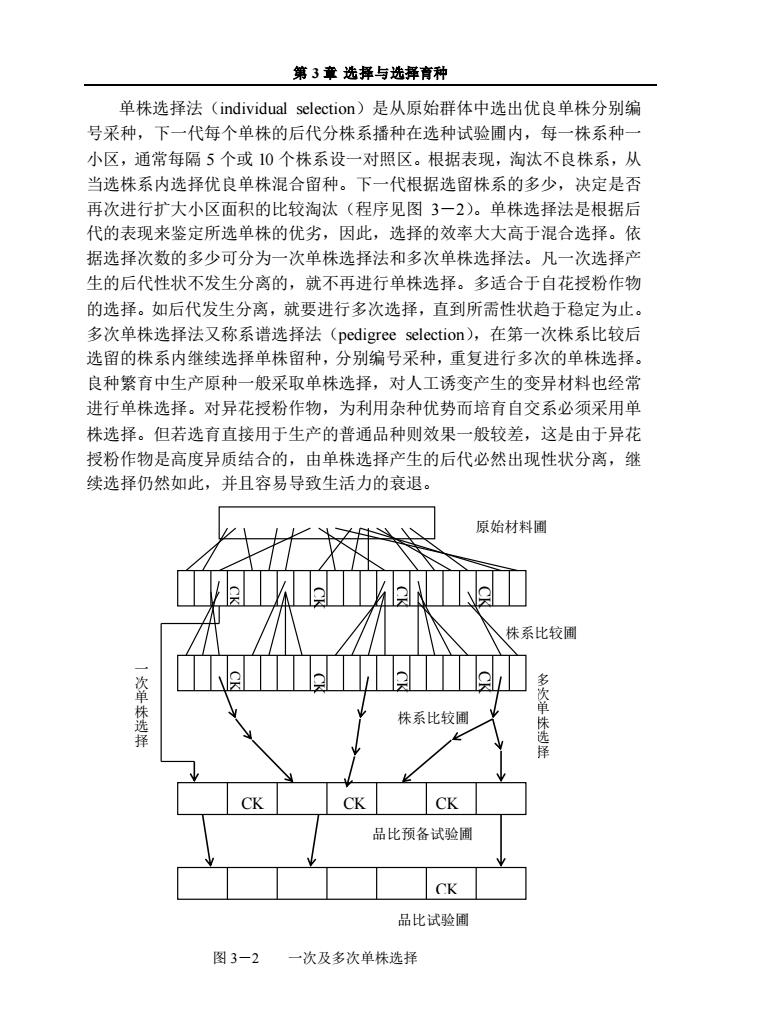
第3章选择与选择育种 单株选择法(individual selection)是从原始群体中选出优良单株分别编 号采种,下一代每个单株的后代分株系播种在选种试验圃内,每一株系种 小区,通常每隔5个或10个株系设一对照区。根据表现,淘汰不良株系,从 当选株系内选择优良单株混合留种。下一代根据选留株系的多少,决定是否 再次进行扩大小区面积的比较淘汰(程序见图3一2)。单株选择法是根据后 代的表现来鉴定所选单株的优劣,因此,选择的效率大大高于混合选择。依 据选择次数的多少可分为一次单株选择法和多次单株选择法。凡一次选择产 生的后代性状不发生分离的,就不再进行单株选择。多适合于自花授粉作物 的选择。如后代发生分离,就要进行多次选择,直到所需性状趋于稳定为止。 多次单株选择法又称系谱选择法(pedigree selection),在第一次株系比较后 选留的株系内继续选择单株留种,分别编号采种,重复进行多次的单株选择。 良种繁育中生产原种一般采取单株选择,对人工诱变产生的变异材料也经常 进行单株选择。对异花授粉作物,为利用杂种优势而培育自交系必须采用单 株选择。但若选育直接用于生产的普通品种则效果一般较差,这是由于异花 授粉作物是高度异质结合的,由单株选择产生的后代必然出现性状分离,继 续选择仍然如此,并且容易导致生活力的衰退。 原始材料圃 株系比较圃 次单株选择 株系比较圃 多次单株选择 CK CK CK 品比预备试验圃 CK 品比试验圃 图3-2 一次及多次单株选择
第 3 章 选择与选择育种 单株选择法(individual selection)是从原始群体中选出优良单株分别编 号采种,下一代每个单株的后代分株系播种在选种试验圃内,每一株系种一 小区,通常每隔 5 个或 10 个株系设一对照区。根据表现,淘汰不良株系,从 当选株系内选择优良单株混合留种。下一代根据选留株系的多少,决定是否 再次进行扩大小区面积的比较淘汰(程序见图 3-2)。单株选择法是根据后 代的表现来鉴定所选单株的优劣,因此,选择的效率大大高于混合选择。依 据选择次数的多少可分为一次单株选择法和多次单株选择法。凡一次选择产 生的后代性状不发生分离的,就不再进行单株选择。多适合于自花授粉作物 的选择。如后代发生分离,就要进行多次选择,直到所需性状趋于稳定为止。 多次单株选择法又称系谱选择法(pedigree selection),在第一次株系比较后 选留的株系内继续选择单株留种,分别编号采种,重复进行多次的单株选择。 良种繁育中生产原种一般采取单株选择,对人工诱变产生的变异材料也经常 进行单株选择。对异花授粉作物,为利用杂种优势而培育自交系必须采用单 株选择。但若选育直接用于生产的普通品种则效果一般较差,这是由于异花 授粉作物是高度异质结合的,由单株选择产生的后代必然出现性状分离,继 续选择仍然如此,并且容易导致生活力的衰退。 图 3-2 一次及多次单株选择 CKCKCK CK 原始材料圃 多 次 单 株 选 择 一 次 单 株 选 择 株系比较圃 品比试验圃 品比预备试验圃 CK CK CK CK CK CK 株系比较圃 CK CK CK CK

第3章选择与选择育种 3.2.3.3根据选择方式分:直接选择和间接选择 直接选择法(direct selection)是指对需要改进的性状本身进行直接的选 择,它对质量性状的选择效果往往较高,在育种过程中经常采用。但当目标 性状是数量性状并且遗传力较低时,如产量等经济性状,直接选择效果往往 较差。由于作物的各性状之间存在着不同程度的相关,对一个性状的选择势 必影响到另一个性状的遗传效果。这样,就可以从另一些在基因型上与这些 经济性状密切相关而遗传力又较高的性状进行选择,从而得到有效的结果。 这种通过选择相关性状而达到提高选择效果而改良目标性状的选择方法称为 间接选择法(indirect selection)。 间接选择还可以用来对那些目标性状表现较晚的材料提早进行相关选 择,这对于多年生果树经济性状的选择有着特别的意义,可以加速选择的进 程,并且使后期选择的工作量大大减少。例如,在樱桃实生选种中可以早在 一年生苗期根据叶基腺体大小和果实大小的相关,淘汰大量未来小果型的单 株。再如桃早熟基因(D)与果肉红色基因(B2)之间表现紧密连锁,实生 苗在秋季时叶片呈现部分红色者与未来果实成熟早亦存在密切相关。另外, 对大量个体直接选择有困难时,可先通过间接选择法淘汰大量无希望的个体, 再对留下的少量个体进行间接选择。如进行实生矮化砧选种时,可根据根皮 率与矮化性能的相关,淘汰大量单株,留下根皮率高的单株作直接选择。然 而,目前有关多年生木本作物这方面的研究还有一定的局限性,有待于进 步研究和发现选择效果较好的间接选择性状。 3.2.3.4根据选择的层次分:形态选择、选择同工酶标记 选择和分子标记选择 在长期以来的植物育种实践中,选择大多是根据植株或群体的形态性状 进行,选择的依据是根据对形态性状及相关形态性状的鉴定结果。遗传标记 是生物分类学、育种学、遗传学和物种进化学等研究的主要技术指标之一。 直到十多年前,植物遗传和育种中所用的标记大多数是基于形态性状的标记, 如开花期、座果节位、果重、果数及株高等,这类标记直观且容易鉴定,但标 记数量有限,容易受到作物栽培环境的影响。而且,要找到与育种目标性状相 关的形态学标记往往上有一定的难度。 同工酶是植物在长期进化过程中基因突变分化和自然适应的结果,酶谱的 差异能在一定程度上反映某些基因和性状表现的差异。从五十年代末开始,特 别是在过去二三十年里同工酶标记得到广泛的发展和应用,其分析手段较为简
第 3 章 选择与选择育种 3.2.3.3 根据选择方式分:直接选择和间接选择 直接选择法(direct selection)是指对需要改进的性状本身进行直接的选 择,它对质量性状的选择效果往往较高,在育种过程中经常采用。但当目标 性状是数量性状并且遗传力较低时,如产量等经济性状,直接选择效果往往 较差。由于作物的各性状之间存在着不同程度的相关,对一个性状的选择势 必影响到另一个性状的遗传效果。这样,就可以从另一些在基因型上与这些 经济性状密切相关而遗传力又较高的性状进行选择,从而得到有效的结果。 这种通过选择相关性状而达到提高选择效果而改良目标性状的选择方法称为 间接选择法(indirect selection)。 间接选择还可以用来对那些目标性状表现较晚的材料提早进行相关选 择,这对于多年生果树经济性状的选择有着特别的意义,可以加速选择的进 程,并且使后期选择的工作量大大减少。例如,在樱桃实生选种中可以早在 一年生苗期根据叶基腺体大小和果实大小的相关,淘汰大量未来小果型的单 株。再如桃早熟基因(D)与果肉红色基因(B2)之间表现紧密连锁,实生 苗在秋季时叶片呈现部分红色者与未来果实成熟早亦存在密切相关。另外, 对大量个体直接选择有困难时,可先通过间接选择法淘汰大量无希望的个体, 再对留下的少量个体进行间接选择。如进行实生矮化砧选种时,可根据根皮 率与矮化性能的相关,淘汰大量单株,留下根皮率高的单株作直接选择。然 而,目前有关多年生木本作物这方面的研究还有一定的局限性,有待于进一 步研究和发现选择效果较好的间接选择性状。 3.2.3.4 根据选择的层次分:形态选择、选择同工酶标记 选择和分子标记选择 在长期以来的植物育种实践中,选择大多是根据植株或群体的形态性状 进行,选择的依据是根据对形态性状及相关形态性状的鉴定结果。遗传标记 是生物分类学、育种学、遗传学和物种进化学等研究的主要技术指标之一。 直到十多年前,植物遗传和育种中所用的标记大多数是基于形态性状的标记, 如开花期、座果节位、果重、果数及株高等,这类标记直观且容易鉴定,但标 记数量有限,容易受到作物栽培环境的影响。而且,要找到与育种目标性状相 关的形态学标记往往上有一定的难度。 同工酶是植物在长期进化过程中基因突变分化和自然适应的结果,酶谱的 差异能在一定程度上反映某些基因和性状表现的差异。从五十年代末开始,特 别是在过去二三十年里同工酶标记得到广泛的发展和应用,其分析手段较为简

第3章选择与选择育种 单,适合较大群体的遗传分析,在植物遗传育种和发育生理方面研究和应用最 多的同工酶是过氧化物酶、酯酶、过氧化氢酶、淀粉酶、亮氨酸氨肽酶、脱氢 酶、磷酸酯酶、磷酸化酶、氨基转移酶、多酚氧化酶和细胞色素氧化酶。但与 形态标记及以后发展起来的分子标记相比,能够利用的同工酶系统的数量非常 有限。同工酶标记也容易受到作物栽培环境及发育阶段的影响。 近年来,分子生物学技术的快速发展为植物遗传育种提供了基于DNA多 态性的分子标记,较形态标记及同工酶标记有明显的优越性:分子标记是遗传 物质一DNA最根本上差异的反映,它几乎不受作物发育环境的影响:大多 数分子标记是共显性的,对隐性农艺性状的选择十分便利:基因组变异极其丰 富,分子标记的数量几乎是无限的:在发育的不同阶段,不同组织的DNA都 可用于标记分析,使得对植株基因型的早期选择成为可能,这一点对于生殖周 期较长的果树品种的选育有着特别的意义。关于分子标记育种将在生物技术育 种一章作较为详尽的介绍。 3.3遗传力与选择效果 一般将作物的性状分为质量性状和数量性状。质量性状,如抗病性、花色 等性状,受单基因或少数经验控制,受环境的影响较小,后代中不同类型容易 区分选择,选择容易获得较高的选择效果。而数量性状,如产量、品质等大部 分经济性状,受多基因控制,性状变异连续,不能分成明显的类型,并且容易 受环境影响,影响选择效果的因素较多。以下就数量性状的选择效果及影响因 素作以介绍。 3.3.1遗传力与遗传进展 对作物数量性状而言,变异既受基因型的影响,也受环境的影响。一个性 状的表型变异包括遗传变异部分和环境变异部分,即Vp=VG十VE。其中VG 在Vp中所占的比重反映了植株对该性状的遗传能力大小,用遗传力()表 示。遗传方差又由加性方差(VD)、显性方差(VH)和上位性方差(V)组成, 后两部分又称为非加性方差。因显性方差随世代数的增加而递减,在纯合状态 时要消失,不能固定。因此,根据加性方差所占的比重大小更能反映亲子间的 相似程度。这样,遗传力又可进一步分为广义遗传力(hg)和狭义遗传力(、2)。 用以下公式表示: he?=V'G VG ×100= ×100 VP VG+VE
第 3 章 选择与选择育种 单,适合较大群体的遗传分析,在植物遗传育种和发育生理方面研究和应用最 多的同工酶是过氧化物酶、酯酶、过氧化氢酶、淀粉酶、亮氨酸氨肽酶、脱氢 酶、磷酸酯酶、磷酸化酶、氨基转移酶、多酚氧化酶和细胞色素氧化酶。但与 形态标记及以后发展起来的分子标记相比,能够利用的同工酶系统的数量非常 有限。同工酶标记也容易受到作物栽培环境及发育阶段的影响。 近年来,分子生物学技术的快速发展为植物遗传育种提供了基于 DNA 多 态性的分子标记,较形态标记及同工酶标记有明显的优越性:分子标记是遗传 物质 DNA 最根本上差异的反映,它几乎不受作物发育环境的影响;大多 数分子标记是共显性的,对隐性农艺性状的选择十分便利;基因组变异极其丰 富,分子标记的数量几乎是无限的;在发育的不同阶段,不同组织的 DNA 都 可用于标记分析,使得对植株基因型的早期选择成为可能,这一点对于生殖周 期较长的果树品种的选育有着特别的意义。关于分子标记育种将在生物技术育 种一章作较为详尽的介绍。 3.3 遗传力与选择效果 一般将作物的性状分为质量性状和数量性状。质量性状,如抗病性、花色 等性状,受单基因或少数经验控制,受环境的影响较小,后代中不同类型容易 区分选择,选择容易获得较高的选择效果。而数量性状,如产量、品质等大部 分经济性状,受多基因控制,性状变异连续,不能分成明显的类型,并且容易 受环境影响,影响选择效果的因素较多。以下就数量性状的选择效果及影响因 素作以介绍。 3.3.1 遗传力与遗传进展 对作物数量性状而言,变异既受基因型的影响,也受环境的影响。一个性 状的表型变异包括遗传变异部分和环境变异部分,即 VP=VG+VE。其中 VG 在 VP 中所占的比重反映了植株对该性状的遗传能力大小,用遗传力(h 2)表 示。遗传方差又由加性方差(VD)、显性方差(VH)和上位性方差(VI)组成, 后两部分又称为非加性方差。因显性方差随世代数的增加而递减,在纯合状态 时要消失,不能固定。因此,根据加性方差所占的比重大小更能反映亲子间的 相似程度。这样,遗传力又可进一步分为广义遗传力(hB 2)和狭义遗传力(hN 2)。 用以下公式表示: hB 2= VP VG ×100= VG VE VG + ×100

第3章选择与选择育种 hN=VD VD ×100= ×100 VP VG+VE 在一定的试验设计,可以对某一群体的上述各项参数进行估算,并计算出 广义遗传力或狭义遗传力。广义遗传力仅仅表示了一个性状遗传因素和环境因 素影响的相对重要性:狭义遗传力代表了总变异中归属于基因的平均效应部 分,反映了亲子间的相似程度,它对于后代表型的预测有重要意义。 对某一数量性状进行选择时,入选群体的平均值将与原始群体平均值产生 一定的离差,即选择差(i,也有用S表示)。例如,在番茄平均果重为150克 的群体中选择果重较高的植株,入选植株的平均果重如为200克,则选择差 =200-150=50克。 选择差在一定程度上反映了选择的效果,但并不能全部遗传给后代。其子 代的平均值通常在原始群体平均值和入选群体平均值之间。这些入选亲本后代 构成群体的平均值与上述原始群体平均值之差用遗传进度(△G,或GS)表示, 也有人称其为遗传获得量、选择响应或选择反应,它反映了选择效果的大小。 如果上例番茄入选群体的后代的平均果重为180克,则遗传进度△G=180一150 =30克。 子代平均值对入选群体平均值的差异倾向程度,决定于该性状遗传力() 的大小。h愈接近1,则子代平均值就愈接近于入选群体的平均值,选择效果 愈好:反之,h2愈接近于0,则子代平均值趋向于原始群体的平均值,选择效 果愈差。在育种实践中可以根据遗传进度与选择差的比值来估计遗传力的大 小。这样估算的遗传力又被称为现实遗传力(h取2),用下列公式表示: hR2=△G i×100 在上例番茄群体中,可以估算出h取2为60%。这是从已经实现的选择的结 果来估算遗传力的方法,估算时应该注意前后代的环境条件应尽可能的一致。 这种遗传力对于选择的有效性提供了一种很好的经验公式。 在育种过程中,经常将上述公式反过来估算预期的遗传进度。遗传进度的 大小反映了选择效果的好坏。这时通常用其它方法估算的狭义遗传力或广义遗 传力来代替现实遗传力,公式为: △G=h2i 为了进一步比较不同群体或性状的选择效果,需要将选择差标准化,这 种标准化后的选择差称为选择强度(K),即以标准差(6)为单位的选择差, 公式为:
第 3 章 选择与选择育种 hN 2= VP VD ×100= VG VE VD + ×100 在一定的试验设计,可以对某一群体的上述各项参数进行估算,并计算出 广义遗传力或狭义遗传力。广义遗传力仅仅表示了一个性状遗传因素和环境因 素影响的相对重要性;狭义遗传力代表了总变异中归属于基因的平均效应部 分,反映了亲子间的相似程度,它对于后代表型的预测有重要意义。 对某一数量性状进行选择时,入选群体的平均值将与原始群体平均值产生 一定的离差,即选择差(i,也有用 S 表示)。例如,在番茄平均果重为 150 克 的群体中选择果重较高的植株,入选植株的平均果重如为 200 克,则选择差 i =200-150=50 克。 选择差在一定程度上反映了选择的效果,但并不能全部遗传给后代。其子 代的平均值通常在原始群体平均值和入选群体平均值之间。这些入选亲本后代 构成群体的平均值与上述原始群体平均值之差用遗传进度(ΔG,或 GS)表示, 也有人称其为遗传获得量、选择响应或选择反应,它反映了选择效果的大小。 如果上例番茄入选群体的后代的平均果重为180克,则遗传进度ΔG=180-150 =30 克。 子代平均值对入选群体平均值的差异倾向程度,决定于该性状遗传力(h 2) 的大小。h 2 愈接近 1,则子代平均值就愈接近于入选群体的平均值,选择效果 愈好;反之,h 2 愈接近于 0,则子代平均值趋向于原始群体的平均值,选择效 果愈差。在育种实践中可以根据遗传进度与选择差的比值来估计遗传力的大 小。这样估算的遗传力又被称为现实遗传力(hR 2),用下列公式表示: hR 2=ΔG i×100 在上例番茄群体中,可以估算出 hR 2 为 60%。这是从已经实现的选择的结 果来估算遗传力的方法,估算时应该注意前后代的环境条件应尽可能的一致。 这种遗传力对于选择的有效性提供了一种很好的经验公式。 在育种过程中,经常将上述公式反过来估算预期的遗传进度。遗传进度的 大小反映了选择效果的好坏。这时通常用其它方法估算的狭义遗传力或广义遗 传力来代替现实遗传力,公式为: ΔG=h 2 i 为了进一步比较不同群体或性状的选择效果,需要将选择差 i 标准化,这 种标准化后的选择差称为选择强度(K),即以标准差(б)为单位的选择差, 公式为:

第3章选择与选择育种 K= 6,或=K6 这样,遗传进度、选择强度和遗传力的关系可以用下式表示: △G=h2-K6p 也就是说,选择效果的好坏与性状的标准差、遗传力和选择强度成正比。 当群体表型值为正态分布时,选择强度取决于入选率q的大小,公式为: K-Z/P 可算得入选率q和选择强度K的对应值(见表3一1)。(Z表示正态分布 曲线右尾入选百分数面积在横坐标截点处的高)。 表3一1选择百分数q与选择强度K的关系 q(%) 0.01 0.05 0.1 0.5 1 2 3 4 5 6 7 K 4.0 3.6 3.4 2.9 2.672.422.272.152.061.981.92 q(%) 8 9 10 20 30 40 50 60 70 80 90 K 1.86 1.801.75 1.40 1.16 0.97 0.800.64 0.50 0.350.19 为了便于比较不同群体、不同性状、不同世代的遗传进度,可以用相对遗 传进度来表示,即 △G'=h2K6p 式中X为整个亲代的群体平均值。 仍以上例番茄群体为例,假定在1000株中选择50株果重较大的植株留种, 则入选率为5%,在上表中查的K值为2.06,如果估算出的遗传力为60%,表 型标准差为20克,那么其遗传进度和相对遗传进度分别为 △G=60%×2.06×20=24.72(克) △G=24.72/150=0.165 应用上述公式,对任何群体的任一性状,只要知道它的平均值和标准差, 知道它的遗传力,在理论上就可以预测按某种百分率选择后的选择效果,而不 需要实际度量各个选株的数值。 另外,在进行间接选择时,只要知道两个性状的遗传相关和遗传力(当然, 这时这两个性状相应的表现型方差61和622也已被估算出来),那么通过对一 个性状的选择(如性状1)就可以用下式预测另一个性状(性状2)的相关遗 传进度(△G*)。 △G*=rg12K(6p22h12h2212
第 3 章 选择与选择育种 K=i б,或 i=Kб 这样,遗传进度、选择强度和遗传力的关系可以用下式表示: ΔG=h 2 ·K·бP 也就是说,选择效果的好坏与性状的标准差、遗传力和选择强度成正比。 当群体表型值为正态分布时,选择强度取决于入选率 q 的大小,公式为: K=Z/P 可算得入选率 q 和选择强度 K 的对应值(见表 3-1)。(Z 表示正态分布 曲线右尾入选百分数面积在横坐标截点处的高)。 表 3-1 选择百分数 q 与选择强度 K 的关系 q(%) 0.01 0.05 0.1 0.5 1 2 3 4 5 6 7 K 4.0 3.6 3.4 2.9 2.67 2.42 2.27 2.15 2.06 1.98 1.92 q(%) 8 9 10 20 30 40 50 60 70 80 90 K 1.86 1.80 1.75 1.40 1.16 0.97 0.80 0.64 0.50 0.35 0.19 为了便于比较不同群体、不同性状、不同世代的遗传进度,可以用相对遗 传进度来表示,即 ΔG΄=h 2 ·K·бP X 式中 X 为整个亲代的群体平均值。 仍以上例番茄群体为例,假定在1000株中选择50株果重较大的植株留种, 则入选率为 5%,在上表中查的 K 值为 2.06,如果估算出的遗传力为 60%,表 型标准差为 20 克,那么其遗传进度和相对遗传进度分别为 ΔG=60%×2.06×20=24.72(克) ΔG΄=24.72/150=0.165 应用上述公式,对任何群体的任一性状,只要知道它的平均值和标准差, 知道它的遗传力,在理论上就可以预测按某种百分率选择后的选择效果,而不 需要实际度量各个选株的数值。 另外,在进行间接选择时,只要知道两个性状的遗传相关和遗传力(当然, 这时这两个性状相应的表现型方差 бP1 和 бP22 也已被估算出来),那么通过对一 个性状的选择(如性状 1)就可以用下式预测另一个性状(性状 2)的相关遗 传进度(ΔG*)。 ΔG*=rg12·K· (бP22h1 2h2 2 ) 1/2