
第四章 芽变育种 芽变($port)来源于体细胞自然发生遗传物质的变异。变异的体细胞发生于芽的分生细 胞中或经分裂发育进入芽的分生组织,就形成变异芽。当变异的芽萌发成枝条,乃至开花或 结果以后,表现出与原品种的性状有明显差异时,才被发现,所以芽变总是以“枝变”的形 式表现。这种变异的枝芽有时在被人们发现前,己被无意识地用于无性繁殖,在长成新的植 株时才被首次发现。这种变异植株称之为“株变”。芽变育种指对由芽变发生的变异进行选 择,从而培育出新品种的一种育种方法。在园艺作物的营养系品种内,除由遗传物质变异而 发生变异外,还普遍存在着由种种环境条件(如砧木,施肥制度,果园地貌,土壤,紫外线 等各种气象因素,以及其他一系列栽培措施的影响)而造成的不能遗传的彷惶变异,又称饰 变。因此,芽变选种的一个首要问题就是正确区分这种变异,选出真正的优良芽变。 芽变现象普遍存在于自然界,我国古代劳动人民很早就发现芽变现象,1031年欧阳修在 “洛阳牡丹记”中记述了牡丹的多种芽变:1059年宋朝蔡襄《荔枝谱》中就有荔枝芽变品 种的记载。据A.D.Shamel及C.S.Pomerey 19936年调查,在22种果树中,仅果实及叶 片的芽变就发现987个。因此,可见芽变育种,己成为无性繁殖作物产生新变异无限丰富的 源泉,不仅直接可以从中选育出优良的新品种,又可不断丰富现有的种质资源库,给杂交育 种提供新的种质资源,是选育新品种的一种简易而有效的方法。芽变育种的突出优点是可对 优良品种的个别缺点进行修缮,同时又能基本上保持其原有的综合性状。这在园艺作物育种 中得到广泛的应用,尤其明显的例子是元帅系苹果由芽变育种而实现的品种演化。 元帅苹果是1880年在美国发现的偶然实生树,1895年定名推广:20世纪20年代,从 元帅中选出了色泽优于元帅的红星和红冠等第二代品种:40~50年代,选出了具有浓红短 枝型特点的新红星、超红和艳红等第三代品种:60~70年代,发现了外观和风味、品质更 优良的魁红、首红和银红等第四代品种:70年代以后,又出现一批着色更早、色泽浓红的 短枝型芽变系俄矮2号和皇家矮红等第五代品种,从而使元帅系苹果不断得到改良和提高。 目前,元帅系芽变品种己达180多个,形成了一个世界范围的庞大芽变品种群。再如日本的 苹果品种富士,于1962年进行种苗登录后,发展缓慢,但自70年代选出一批着色好的芽变 品系后,发展迅速:至1984年其面积和产量都已跃居日本苹果栽培的第一位,使日本的苹 果栽培品种组成发生了很大的变化。在观赏类植物中芽变育种也有重要意义。例如牡丹花的 花型进化,先由单瓣类发展到半重瓣类(即千层类),以至最后形成重瓣类(即楼子类),即 是一个芽变育种的过程。在草本植物中应用不如木本植物明显,由于其生长周期短,芽变单 株不易发现,而且受环境影响因素大,饰变成份较多,所以在草本(如多数蔬菜、花卉、部 分果树)园艺作物中芽变育种应用居于次要地位。总之,芽变育种不仅在历史上起到品种改 良的作用,而且特别是近代,在国内外很受重视。因为芽变育种方法简便,收效较快,易为 从事生产人员所掌握。我国地大物博,园艺作物栽培历史悠久,资源丰富,为开展芽变育种 提供了大量的资源。我们要充分利用这一有利条件,采用专业研究机构与群众选种相结合的 方法,持续深入地开展芽变育种工作,不断地育出更好的新品种。本章将主要针对应用性较 强的多年生园艺作物的芽变育种作一介绍
1 第四章 芽变育种 芽变(sport)来源于体细胞自然发生遗传物质的变异。变异的体细胞发生于芽的分生细 胞中或经分裂发育进入芽的分生组织,就形成变异芽。当变异的芽萌发成枝条,乃至开花或 结果以后,表现出与原品种的性状有明显差异时,才被发现,所以芽变总是以“枝变”的形 式表现。这种变异的枝芽有时在被人们发现前,已被无意识地用于无性繁殖,在长成新的植 株时才被首次发现。这种变异植株称之为“株变”。芽变育种指对由芽变发生的变异进行选 择,从而培育出新品种的一种育种方法。在园艺作物的营养系品种内,除由遗传物质变异而 发生变异外,还普遍存在着由种种环境条件(如砧木,施肥制度,果园地貌,土壤,紫外线 等各种气象因素,以及其他一系列栽培措施的影响)而造成的不能遗传的彷惶变异,又称饰 变。因此,芽变选种的一个首要问题就是正确区分这种变异,选出真正的优良芽变。 芽变现象普遍存在于自然界,我国古代劳动人民很早就发现芽变现象,l031 年欧阳修在 “洛阳牡丹记”中记述了牡丹的多种芽变;1059 年宋朝蔡襄《荔枝谱》中就有荔枝芽变品 种的记载。据 A.D.Shamel 及 C.S.Pomerey l936 年调查,在 22 种果树中,仅果实及叶 片的芽变就发现 987 个。因此,可见芽变育种,已成为无性繁殖作物产生新变异无限丰富的 源泉,不仅直接可以从中选育出优良的新品种,又可不断丰富现有的种质资源库,给杂交育 种提供新的种质资源,是选育新品种的一种简易而有效的方法。芽变育种的突出优点是可对 优良品种的个别缺点进行修缮,同时又能基本上保持其原有的综合性状。这在园艺作物育种 中得到广泛的应用,尤其明显的例子是元帅系苹果由芽变育种而实现的品种演化。 元帅苹果是 1880 年在美国发现的偶然实生树,1895 年定名推广;20 世纪 20 年代,从 元帅中选出了色泽优于元帅的红星和红冠等第二代品种;40~50 年代,选出了具有浓红短 枝型特点的新红星、超红和艳红等第三代品种;60~70 年代,发现了外观和风味、品质更 优良的魁红、首红和银红等第四代品种;70 年代以后,又出现一批着色更早、色泽浓红的 短枝型芽变系俄矮 2 号和皇家矮红等第五代品种,从而使元帅系苹果不断得到改良和提高。 目前,元帅系芽变品种已达 180 多个,形成了一个世界范围的庞大芽变品种群。再如日本的 苹果品种富士,于 1962 年进行种苗登录后,发展缓慢,但自 70 年代选出一批着色好的芽变 品系后,发展迅速;至 1984 年其面积和产量都已跃居日本苹果栽培的第一位,使日本的苹 果栽培品种组成发生了很大的变化。在观赏类植物中芽变育种也有重要意义。例如牡丹花的 花型进化,先由单瓣类发展到半重瓣类(即千层类),以至最后形成重瓣类(即楼子类),即 是一个芽变育种的过程。在草本植物中应用不如木本植物明显,由于其生长周期短,芽变单 株不易发现,而且受环境影响因素大,饰变成份较多,所以在草本(如多数蔬菜、花卉、部 分果树)园艺作物中芽变育种应用居于次要地位。总之,芽变育种不仅在历史上起到品种改 良的作用,而且特别是近代,在国内外很受重视。因为芽变育种方法简便,收效较快,易为 从事生产人员所掌握。我国地大物博,园艺作物栽培历史悠久,资源丰富,为开展芽变育种 提供了大量的资源。我们要充分利用这一有利条件,采用专业研究机构与群众选种相结合的 方法,持续深入地开展芽变育种工作,不断地育出更好的新品种。本章将主要针对应用性较 强的多年生园艺作物的芽变育种作一介绍

第一节芽变的遗传学基础 一、芽变的类型及特点 (一)芽变遗传类型 芽变是遗传物质的变异,主要表现为染色体数量变异、染色体结构变异、基因突变和核 外突变四种类型,说明了遗传物质不同层次的变异结果。 1.染色体数目变异,包括多倍体、单倍体及非整倍体,主要是多倍体的突变,其往往 表现各种器官的巨大性。葡萄无核白,为四倍体芽变:玫瑰香的四倍体芽变“大玫瑰香”, 其果穗、果粒比玫瑰香明显增大。 2.染色体结构变异,包括易位,倒位,重复及缺失,是由于染色体结构发生变异而造 成基因线性顺序发生变化,从而使有关性状发生变异。这类变异可以通过扦插,嫁接等无性 繁殖方法保存下来。 3.基因突变,通常是由一个基因突变形成的,同时几个基因发生突变是极稀少的,如 苹果短枝型芽变,不仅是枝条节间变短,而且变粗,树冠矮化,早果丰产。这些性状之间可 能是一因多效关系,而并非是几个基因同时发生突变。 4.核外突变,指的是细胞质中的遗传物质发生突变。有些性状是细胞质基因控制的, 特别是母性遗传效应比较明显的作物,在育性、性分化、叶绿素形成、植株高度和生活力等 方面性状发生了变异,有可能是细胞质基因发生突变的结果: (二)芽变的特点 对于任何发生在性细胞上的突变通常都有可能发生于体细胞。遗传学中所总结的突变规 律基本适用于芽变,如突变的可逆性,正突变频率大于逆突变:突变的多向性产生复等位基 因:突变的一般有害性等。这里主要介绍与芽变育种有关的体细胞突变的一些特点。 1.芽变的嵌合性体细胞突变最初仅发生于个别细胞,就发生突变的个体、器官或组 织而言,它是由突变与未突变细胞组成的嵌合体(chimera)。体细胞在细胞分裂、发育过程 中由异型细胞间的竞争和选择的作用而转化成突变芽、枝、植株系。芽变育种就是促进优良 的突变体细胞实现这种转化,从而育成在无性繁殖中能稳定遗传的芽变品种。有些观赏植物, 如花卉的“二乔”、“跳枝”类型,竹类的黄金间碧玉类型要求某种程度的异型嵌合状态。有 时芽变选种很难使突变体达到100%同型化,所以不少芽变品种会偶然出现原品种性状而不 够稳定,应持续选择,提高其同型化程度。 2.芽变的平行性瓦维洛夫(1926)在资源考察中发现相近植物的属中存在遗传变异 平行规律。它完全适用于体细胞突变,且对芽变育种有着重要的指导意义。如在桃的芽变中 曾经出现过重瓣、红花、花粉不育、粘核、垂枝、短枝型、早熟等芽变。人们就能有把握地 期待在李亚科的其它属、种如杏、梅、樱桃中出现平行的芽变类型,甚至能预测梨亚科、蔷 薇亚科的不同树种像苹果、蔷薇会发生除黏核以外的其余所有芽变类型。在种内品种间和同 品种的植株间这种平行性芽变机率会更高一些。如自美国50年代从元帅系苹果中选育出短 枝型芽变品种新红星以来,中国各地不仅从元帅系品种,而且从金冠、富士、国光、白龙等 苹果品种中陆续选育出系列短枝型新品种。 3.芽变的多样性在“形态特征”上体现的最为突出,包括突变部位的多样性,突变
2 第一节 芽变的遗传学基础 一、芽变的类型及特点 (一)芽变遗传类型 芽变是遗传物质的变异,主要表现为染色体数量变异、染色体结构变异、基因突变和核 外突变四种类型,说明了遗传物质不同层次的变异结果。 1.染色体数目变异,包括多倍体、单倍体及非整倍体,主要是多倍体的突变,其往往 表现各种器官的巨大性。葡萄无核白,为四倍体芽变;玫瑰香的四倍体芽变“大玫瑰香”, 其果穗、果粒比玫瑰香明显增大。 2.染色体结构变异,包括易位,倒位,重复及缺失,是由于染色体结构发生变异而造 成基因线性顺序发生变化,从而使有关性状发生变异。这类变异可以通过扦插,嫁接等无性 繁殖方法保存下来。 3.基因突变,通常是由一个基因突变形成的,同时几个基因发生突变是极稀少的,如 苹果短枝型芽变,不仅是枝条节间变短,而且变粗,树冠矮化,早果丰产。这些性状之间可 能是一因多效关系,而并非是几个基因同时发生突变。 4.核外突变,指的是细胞质中的遗传物质发生突变。有些性状是细胞质基因控制的, 特别是母性遗传效应比较明显的作物,在育性、性分化、叶绿素形成、植株高度和生活力等 方面性状发生了变异,有可能是细胞质基因发生突变的结果。 (二)芽变的特点 对于任何发生在性细胞上的突变通常都有可能发生于体细胞。遗传学中所总结的突变规 律基本适用于芽变,如突变的可逆性,正突变频率大于逆突变;突变的多向性产生复等位基 因;突变的一般有害性等。这里主要介绍与芽变育种有关的体细胞突变的一些特点。 1.芽变的嵌合性 体细胞突变最初仅发生于个别细胞,就发生突变的个体、器官或组 织而言,它是由突变与未突变细胞组成的嵌合体(chimera)。体细胞在细胞分裂、发育过程 中由异型细胞间的竞争和选择的作用而转化成突变芽、枝、植株系。芽变育种就是促进优良 的突变体细胞实现这种转化,从而育成在无性繁殖中能稳定遗传的芽变品种。有些观赏植物, 如花卉的“二乔”、“跳枝”类型,竹类的黄金间碧玉类型要求某种程度的异型嵌合状态。有 时芽变选种很难使突变体达到 100%同型化,所以不少芽变品种会偶然出现原品种性状而不 够稳定,应持续选择,提高其同型化程度。 2.芽变的平行性 瓦维洛夫(1926)在资源考察中发现相近植物的属中存在遗传变异 平行规律。它完全适用于体细胞突变,且对芽变育种有着重要的指导意义。如在桃的芽变中 曾经出现过重瓣、红花、花粉不育、粘核、垂枝、短枝型、早熟等芽变。人们就能有把握地 期待在李亚科的其它属、种如杏、梅、樱桃中出现平行的芽变类型,甚至能预测梨亚科、蔷 薇亚科的不同树种像苹果、蔷薇会发生除黏核以外的其余所有芽变类型。在种内品种间和同 品种的植株间这种平行性芽变机率会更高一些。如自美国 50 年代从元帅系苹果中选育出短 枝型芽变品种新红星以来,中国各地不仅从元帅系品种,而且从金冠、富士、国光、白龙等 苹果品种中陆续选育出系列短枝型新品种。 3.芽变的多样性 在“形态特征”上体现的最为突出,包括突变部位的多样性,突变
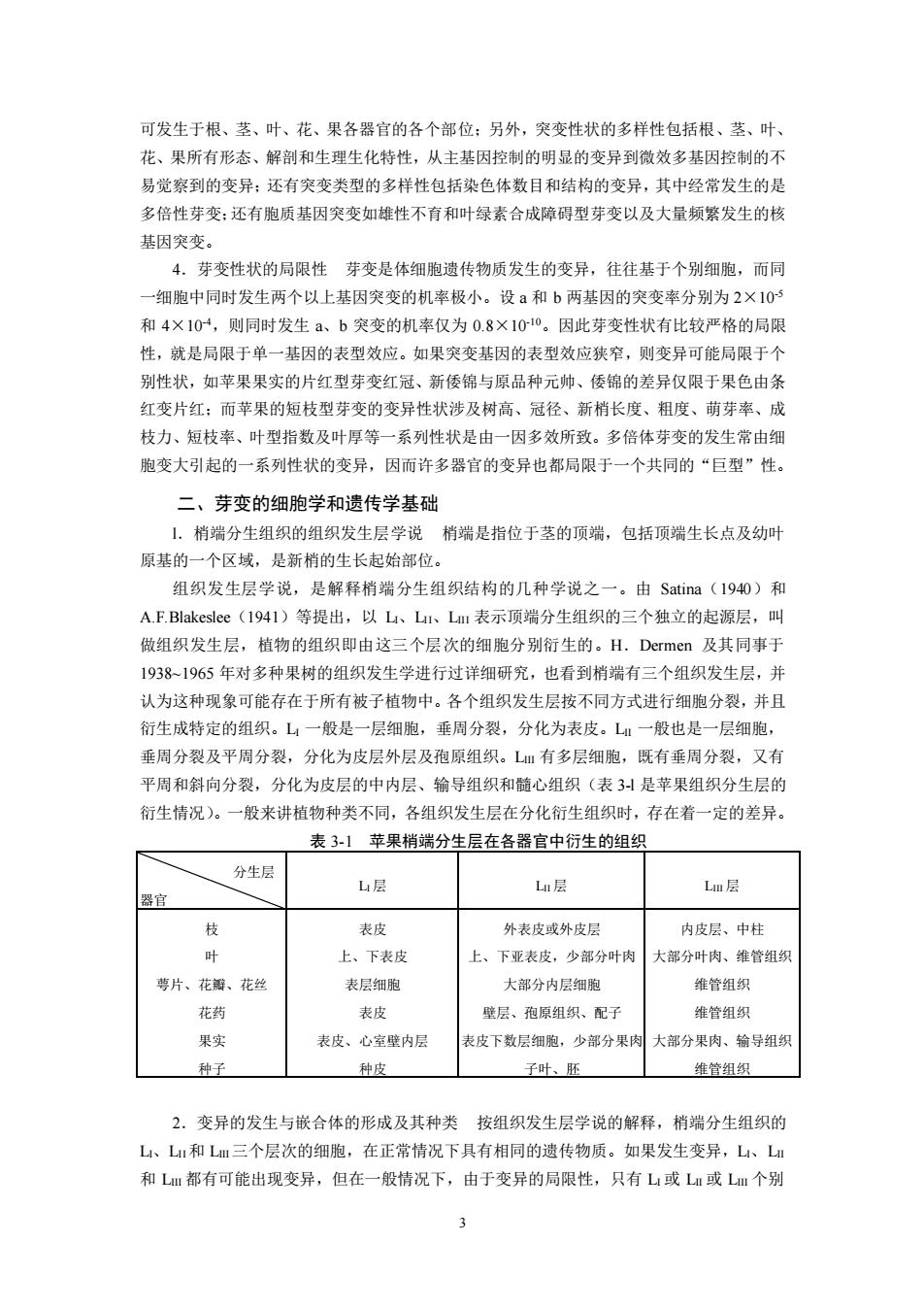
可发生于根、茎、叶、花、果各器官的各个部位;另外,突变性状的多样性包括根、茎、叶、 花、果所有形态、解剖和生理生化特性,从主基因控制的明显的变异到微效多基因控制的不 易觉察到的变异:还有突变类型的多样性包括染色体数目和结构的变异,其中经常发生的是 多倍性芽变:还有胞质基因突变如雄性不育和叶绿素合成障碍型芽变以及大量频繁发生的核 基因突变。 4.芽变性状的局限性芽变是体细胞遗传物质发生的变异,往往基于个别细胞,而同 一细胞中同时发生两个以上基因突变的机率极小。设a和b两基因的突变率分别为2×105 和4×104,则同时发生a、b突变的机率仅为0.8×1010。因此芽变性状有比较严格的局限 性,就是局限于单一基因的表型效应。如果突变基因的表型效应狭窄,则变异可能局限于个 别性状,如苹果果实的片红型芽变红冠、新倭锦与原品种元帅、倭锦的差异仅限于果色由条 红变片红:而苹果的短枝型芽变的变异性状涉及树高、冠径、新梢长度、粗度、萌芽率、成 枝力、短枝率、叶型指数及叶厚等一系列性状是由一因多效所致。多倍体芽变的发生常由细 胞变大引起的一系列性状的变异,因而许多器官的变异也都局限于一个共同的“巨型”性。 二、芽变的细胞学和遗传学基础 L.梢端分生组织的组织发生层学说梢端是指位于茎的顶端,包括顶端生长点及幼叶 原基的一个区域,是新梢的生长起始部位。 组织发生层学说,是解释梢端分生组织结构的几种学说之一。由Satina(1940)和 A.F.Blakeslee(I941)等提出,以L、LuI、Lml表示顶端分生组织的三个独立的起源层,叫 做组织发生层,植物的组织即由这三个层次的细胞分别衍生的。H.Dermen及其同事于 1938~1965年对多种果树的组织发生学进行过详细研究,也看到梢端有三个组织发生层,并 认为这种现象可能存在于所有被子植物中。各个组织发生层按不同方式进行细胞分裂,并且 衍生成特定的组织。L一般是一层细胞,垂周分裂,分化为表皮。L一般也是一层细胞, 垂周分裂及平周分裂,分化为皮层外层及孢原组织。LⅢ有多层细胞,既有垂周分裂,又有 平周和斜向分裂,分化为皮层的中内层、输导组织和髓心组织(表3!是苹果组织分生层的 衍生情况)。一般来讲植物种类不同,各组织发生层在分化衍生组织时,存在着一定的差异。 表3-1苹果梢端分生层在各器官中衍生的组织 分生层 L层 L层 Lm层 器官 枝 表皮 外表皮或外皮层 内皮层、中柱 4 上、下表皮 上、下亚表皮,少部分叶肉 大部分叶肉、维管组织 萼片、花瓣、花丝 表层细胞 大部分内层细胞 维管组织 花药 表皮 壁层、孢原组织、配子 维管组织 果实 表皮、心室壁内层 表皮下数层细胞,少部分果肉大部分果肉、输导组织 种子 种皮 子叶、胚 维管组织 2.变异的发生与嵌合体的形成及其种类按组织发生层学说的解释,梢端分生组织的 L、L!和Lm三个层次的细胞,在正常情况下具有相同的遗传物质。如果发生变异,L、L 和LⅢ都有可能出现变异,但在一般情况下,由于变异的局限性,只有L或L!或Lm个别
3 可发生于根、茎、叶、花、果各器官的各个部位;另外,突变性状的多样性包括根、茎、叶、 花、果所有形态、解剖和生理生化特性,从主基因控制的明显的变异到微效多基因控制的不 易觉察到的变异;还有突变类型的多样性包括染色体数目和结构的变异,其中经常发生的是 多倍性芽变;还有胞质基因突变如雄性不育和叶绿素合成障碍型芽变以及大量频繁发生的核 基因突变。 4.芽变性状的局限性 芽变是体细胞遗传物质发生的变异,往往基于个别细胞,而同 一细胞中同时发生两个以上基因突变的机率极小。设 a 和 b 两基因的突变率分别为 2×10-5 和 4×10-4,则同时发生 a、b 突变的机率仅为 0.8×10-10。因此芽变性状有比较严格的局限 性,就是局限于单一基因的表型效应。如果突变基因的表型效应狭窄,则变异可能局限于个 别性状,如苹果果实的片红型芽变红冠、新倭锦与原品种元帅、倭锦的差异仅限于果色由条 红变片红;而苹果的短枝型芽变的变异性状涉及树高、冠径、新梢长度、粗度、萌芽率、成 枝力、短枝率、叶型指数及叶厚等一系列性状是由一因多效所致。多倍体芽变的发生常由细 胞变大引起的一系列性状的变异,因而许多器官的变异也都局限于一个共同的“巨型”性。 二、芽变的细胞学和遗传学基础 l.梢端分生组织的组织发生层学说 梢端是指位于茎的顶端,包括顶端生长点及幼叶 原基的一个区域,是新梢的生长起始部位。 组织发生层学说,是解释梢端分生组织结构的几种学说之一。由 Satina(1940)和 A.F.Blakeslee(1941)等提出,以 LI、LII、LIII 表示顶端分生组织的三个独立的起源层,叫 做组织发生层,植物的组织即由这三个层次的细胞分别衍生的。H.Dermen 及其同事于 1938~1965 年对多种果树的组织发生学进行过详细研究,也看到梢端有三个组织发生层,并 认为这种现象可能存在于所有被子植物中。各个组织发生层按不同方式进行细胞分裂,并且 衍生成特定的组织。LI 一般是一层细胞,垂周分裂,分化为表皮。LII 一般也是一层细胞, 垂周分裂及平周分裂,分化为皮层外层及孢原组织。LIII 有多层细胞,既有垂周分裂,又有 平周和斜向分裂,分化为皮层的中内层、输导组织和髓心组织(表 3-l 是苹果组织分生层的 衍生情况)。一般来讲植物种类不同,各组织发生层在分化衍生组织时,存在着一定的差异。 表 3-1 苹果梢端分生层在各器官中衍生的组织 分生层 器官 LI 层 LII 层 LIII 层 枝 叶 萼片、花瓣、花丝 花药 果实 种子 表皮 上、下表皮 表层细胞 表皮 表皮、心室壁内层 种皮 外表皮或外皮层 上、下亚表皮,少部分叶肉 大部分内层细胞 壁层、孢原组织、配子 表皮下数层细胞,少部分果肉 子叶、胚 内皮层、中柱 大部分叶肉、维管组织 维管组织 维管组织 大部分果肉、输导组织 维管组织 2.变异的发生与嵌合体的形成及其种类 按组织发生层学说的解释,梢端分生组织的 LI、LII和 LIII三个层次的细胞,在正常情况下具有相同的遗传物质。如果发生变异,LI、LII 和 LIII 都有可能出现变异,但在一般情况下,由于变异的局限性,只有 LI 或 LII 或 LIII 个别

层中个别细胞发生变异,三层同时发生同一变异的可能性,几乎是不存在的。在突变发生之 后,变与未变的细胞成为同时分裂、竞争共存的嵌合体(chimera)。因此,芽变开始发生时 总是以嵌合体的形式出现。由于突变发生的时期早迟、突变细胞在变异发生时所处的位置及 以后在分裂过程中发生的层间取代作用,致使形成的嵌合体有多种结构类型。当突变发生时 间早,梢端正在分裂的细胞数少突变细胞又位于某一组织的最中心处,则突变有可能发育成 层间基因型不同的周缘嵌合体(periclinal chimera)。如果突变发生的时间较晚,梢端正在分 裂的细胞数多,突变细胞的位置又不在中心,则变异细胞只能占据层内的一部分使同一层次 内兼有变和未变的两类细胞,称为扇形嵌合体(sectorial chimera),或叫做部分周缘嵌合体。 图3-1梢端组织发生层由突变形成的嵌合体主要类型示意图 1.内周2.中周3.外周4.内扇5.中扇6.外扇 根据突变细胞所处的层次,周缘嵌合体和扇形嵌合体均包括多种类型结构。为便于表达, 现以original(原始的)一词的第一个字母O代表未变的细胞组织,以mutational(突变的) 一词第一个字母m代表突变的细胞组织,按L-L-LⅢ的层次排列,则周缘嵌合体的结构有 m-0-0,0m-0,0-0-m,m-m-0,0-m-m,m-0-m六种类型:扇形嵌合体的结构类别有0.m-0-0 (0.m表示未变与突变细胞存在于同层内,写在前面的符号表示数量上占优势,下同), 0-0.m-0,0-0-0.m,0.m-0.m-0,0-0.m-0.m,0.m-0-0.m,0m-0.m-0.m等类型(图3-1)。此外, 对于染色体倍数性的芽变,则常用4-2-2,2-4-4,4-48,8-4-4等表示其不同的倍性嵌合类 别。 3.嵌合体的转化与芽变性状的稳定性园艺作物芽变体有些变异性状很稳定,一经改 变,就能在生命周期中长期保持,无论采取何种方法繁殖,都能把变异性状传给后代:有些 变异性状,只能在无性繁殖下保持,一旦有性繁殖就出现分离,或全部恢复为原有类型:还 有一些变异性状,即使无性繁殖也不稳定,一部分后代会失去变异性状,恢复为原类型。例 如,起源于苹果旭的短枝型芽变威赛旭和本迪旭等,不但无性繁殖能保持短枝性状,有性繁 殖也能传给后代。但从总体上看来大部分芽变嵌合体都不是很稳定的,往往会在以后的生长 发育或营养繁殖过程中发生转化。 如图3-2所示,一个扇形嵌合体在发生侧枝时,处于变异扇形面内的芽,萌发后将转化 成具周缘嵌合体的新枝:处于扇形面以外的芽,萌发后将长成非突变枝:而恰巧正处于扇形 嵌合体结构的枝条。由于先端优势、自然伤口或人为短截修剪等因素,往往可使枝条上不同 节位芽具有不均等的萌发成枝的机会,从而使一个原是扇形嵌合体的枝条出现不同情况的转 化。 各种周缘嵌合体芽变,也会在继续生长发育中出现不同变化。例如,一个m-0-0型周缘 嵌合体,可回复到0-0-0型而失去变异特征:也可由原m-m-0型即包括L、L层变异的周
4 层中个别细胞发生变异,三层同时发生同一变异的可能性,几乎是不存在的。在突变发生之 后,变与未变的细胞成为同时分裂、竞争共存的嵌合体(chimera)。因此,芽变开始发生时 总是以嵌合体的形式出现。由于突变发生的时期早迟、突变细胞在变异发生时所处的位置及 以后在分裂过程中发生的层间取代作用,致使形成的嵌合体有多种结构类型。当突变发生时 间早,梢端正在分裂的细胞数少突变细胞又位于某一组织的最中心处,则突变有可能发育成 层间基因型不同的周缘嵌合体(periclinal chimera)。如果突变发生的时间较晚,梢端正在分 裂的细胞数多,突变细胞的位置又不在中心,则变异细胞只能占据层内的一部分使同一层次 内兼有变和未变的两类细胞,称为扇形嵌合体(sectorial chimera),或叫做部分周缘嵌合体。 图 3-1 梢端组织发生层由突变形成的嵌合体主要类型示意图 1.内周 2.中周 3.外周 4.内扇 5.中扇 6.外扇 根据突变细胞所处的层次,周缘嵌合体和扇形嵌合体均包括多种类型结构。为便于表达, 现以 original(原始的)一词的第一个字母 O 代表未变的细胞组织,以 mutational(突变的) 一词第一个字母 m 代表突变的细胞组织,,按 LI-LII-LIII的层次排列,则周缘嵌合体的结构有 m-o-o,o-m-o,o-o-m,m-m-o,o-m-m,m-o-m 六种类型;扇形嵌合体的结构类别有 o.m-o-o (o.m 表示未变与突变细胞存在于同层内,写在前面的符号表示数量上占优势,下同), o-o.m-o,o-o-o.m,o.m-o.m-o,o-o.m-o.m,o.m-o-o.m,o.m-o.m-o.m 等类型(图 3-1)。此外, 对于染色体倍数性的芽变,则常用 4-2-2,2-4-4,4-4-8,8-4-4 等表示其不同的倍性嵌合类 别。 3.嵌合体的转化与芽变性状的稳定性 园艺作物芽变体有些变异性状很稳定,一经改 变,就能在生命周期中长期保持,无论采取何种方法繁殖,都能把变异性状传给后代;有些 变异性状,只能在无性繁殖下保持,一旦有性繁殖就出现分离,或全部恢复为原有类型;还 有一些变异性状,即使无性繁殖也不稳定,一部分后代会失去变异性状,恢复为原类型。例 如,起源于苹果旭的短枝型芽变威赛旭和本迪旭等,不但无性繁殖能保持短枝性状,有性繁 殖也能传给后代。但从总体上看来大部分芽变嵌合体都不是很稳定的,往往会在以后的生长 发育或营养繁殖过程中发生转化。 如图 3-2 所示,一个扇形嵌合体在发生侧枝时,处于变异扇形面内的芽,萌发后将转化 成具周缘嵌合体的新枝;处于扇形面以外的芽,萌发后将长成非突变枝;而恰巧正处于扇形 嵌合体结构的枝条。由于先端优势、自然伤口或人为短截修剪等因素,往往可使枝条上不同 节位芽具有不均等的萌发成枝的机会,从而使一个原是扇形嵌合体的枝条出现不同情况的转 化。 各种周缘嵌合体芽变,也会在继续生长发育中出现不同变化。例如,一个 m-o-o 型周缘 嵌合体,可回复到 o-o-o 型而失去变异特征;也可由原 m-m-o 型即包括 LI、LII层变异的周

缘嵌合体,变成m-m-m型的同型突变体。这种转 化现象是由于突变部分与非突变部分的竞争, 方排挤与取代另一方的结果,叫“层间取代”。有 时外部原因也会造成这种转化,促使组织发生层 的重排而发生变异。如辐射育种时由射线杀死外 层细胞,由冻害等造成不定芽的萌发,均可造成 由深层取代外层。如果深层原来就是与外层不同 的突变体,其结果就会出现新的变异。 4.芽变的遗传效应芽变性状有些能在有性 过程中遗传,有些则只能在无性繁殖时保持稳定, 图3-2嵌合体的自然转化示意图 1.由扇形嵌合体长出的周缘嵌合体枝 这是由于突变发生于不同组织层的缘故。因为决 2.由扇形嵌合体长出的扇形嵌合体枝 定有性过程的孢原组织是由L!层产生,只有当突 3.由扇形嵌合体长出的非嵌合体枝 变包含L层时,才能在有性过程中产生遗传效应。 例如,苹果品种旭产生的两个短枝型芽变类型本迪旭和威赛旭,在杂交育种中都可将短枝性 状传递给后代,当它与金帅杂交时,后代中约有50%属短枝型,而旭的另一个人工诱发短 枝型变异,在与金帅杂交时,后代却未出现短枝型。据分析是前一个自然变异包含L,后 一个诱发变异没有包含L。而L比较单纯,它主要与表皮相关的性状有关系,如茸毛和针 刺的有无。根常由LⅢ深层组织中长出来,所以通过根插,即能得到同质突变体。 对于一对基因控制的性状,突变的发生有四种情况:(1)AA→Aa:(2)Aa→aa:(3) aa→Aa:(4)Aa→AA。在完全显性的情况下,上述(1)和(4)当代不能表现出突变性状, 只有在下一代有性世代的分离中表现出来,而(2)和(3)能在当代显现突变性状。在无性 繁殖情况下,a→Aa的突变性状无需纯化即可得到固定,但自交作物必须通过自交纯化即 可得到固定(自交不亲和性往往是最大的障碍),才能把突变性状稳定下来。在不完全显性 的情况下,或是在基因产生加性效应的情况下,(1)或(4)的突变,能在当代表现。Dommergues 等(1966)报道“处理同质结合的观赏植物种子,所得植株的体细胞突变很少,而处理正在 分离中的种子材料,所得体细胞变异的就多”。S.Broertjes(I967)认为“杂结合的栽培类 型容易发生突变,而开白花的植物极少产生花色的突变”。 第二节芽变育种的方法与程序 一、芽变育种的方法 1.育种目标及时期 芽变育种通常是以原有优良品种为对象,在保持原品种优良性状的基础上,通过选择而 使其个别缺点得到改善,所以育种目标针对性较强。例如,在柑橘的芽变选种中,同样是选 育适于加工糖水橘瓣罐头用品种,现有品种本地早其加工成品的色、香、味、形均极好,育 种的主要目标性状应着重选出无核或少核型:而对现有品种温州蜜柑,由于其本身无籽,芽 变育种的性状应着重于果形、瓣形、汁胞等的加工适应性。 在芽变育种时期,为提高芽变育种的效率,除在整个生长发育期进行细微观察选择外, 应根据育种目标着重抓住目标性状最易发现的时期。例如,对观花植物的芽变育种,应着重
5 图 3-2 嵌合体的自然转化示意图 1.由扇形嵌合体长出的周缘嵌合体枝 2.由扇形嵌合体长出的扇形嵌合体枝 3.由扇形嵌合体长出的非嵌合体枝 缘嵌合体,变成 m-m-m 型的同型突变体。这种转 化现象是由于突变部分与非突变部分的竞争,一 方排挤与取代另一方的结果,叫“层间取代”。有 时外部原因也会造成这种转化,促使组织发生层 的重排而发生变异。如辐射育种时由射线杀死外 层细胞,由冻害等造成不定芽的萌发,均可造成 由深层取代外层。如果深层原来就是与外层不同 的突变体,其结果就会出现新的变异。 4.芽变的遗传效应 芽变性状有些能在有性 过程中遗传,有些则只能在无性繁殖时保持稳定, 这是由于突变发生于不同组织层的缘故。因为决 定有性过程的孢原组织是由 LII层产生,只有当突 变包含 LII层时,才能在有性过程中产生遗传效应。 例如,苹果品种旭产生的两个短枝型芽变类型本迪旭和威赛旭,在杂交育种中都可将短枝性 状传递给后代,当它与金帅杂交时,后代中约有 50%属短枝型,而旭的另一个人工诱发短 枝型变异,在与金帅杂交时,后代却未出现短枝型。据分析是前一个自然变异包含 LII,后 一个诱发变异没有包含 LII。而 LI比较单纯,它主要与表皮相关的性状有关系,如茸毛和针 刺的有无。根常由 LIII深层组织中长出来,所以通过根插,即能得到同质突变体。 对于一对基因控制的性状,突变的发生有四种情况:(1)AA→Aa;(2)Aa→aa;(3) aa→Aa;(4)Aa→AA。在完全显性的情况下,上述(1)和(4)当代不能表现出突变性状, 只有在下一代有性世代的分离中表现出来,而(2)和(3)能在当代显现突变性状。在无性 繁殖情况下,aa→Aa 的突变性状无需纯化即可得到固定,但自交作物必须通过自交纯化即 可得到固定(自交不亲和性往往是最大的障碍),才能把突变性状稳定下来。在不完全显性 的情况下,或是在基因产生加性效应的情况下,(1)或(4)的突变,能在当代表现。Dommergues 等(1966)报道“处理同质结合的观赏植物种子,所得植株的体细胞突变很少,而处理正在 分离中的种子材料,所得体细胞变异的就多”。S.Broertjes(1967)认为“杂结合的栽培类 型容易发生突变,而开白花的植物极少产生花色的突变”。 第二节 芽变育种的方法与程序 一、芽变育种的方法 1.育种目标及时期 芽变育种通常是以原有优良品种为对象,在保持原品种优良性状的基础上,通过选择而 使其个别缺点得到改善,所以育种目标针对性较强。例如,在柑橘的芽变选种中,同样是选 育适于加工糖水橘瓣罐头用品种,现有品种本地早其加工成品的色、香、味、形均极好,育 种的主要目标性状应着重选出无核或少核型;而对现有品种温州蜜柑,由于其本身无籽,芽 变育种的性状应着重于果形、瓣形、汁胞等的加工适应性。 在芽变育种时期,为提高芽变育种的效率,除在整个生长发育期进行细微观察选择外, 应根据育种目标着重抓住目标性状最易发现的时期。例如,对观花植物的芽变育种,应着重