
例上,可以很容易理辉这个问题。当心肌收缩能力不变时,收缩释放的能量是恒定的。如果室内压力升高,表示心肌产生张力消耗的能量相对 增加,用于心肌纤维缩短将相对减少:如果肌肉作等长收缩,释放的能量几乎全部用于产生张力。另一方面,也应该看到,由于动脉压影响了 搏出量,又常常继发地引起一些调节机制的活动。如上述,动脉压的突然升高首先导致搏出量减少,结束造成心室内利余血量增加,充盈量增 加,后者又再引起异长调节机制的调节,搏出量可以恢复正常水平,即通过异长调节可以使动脉压增亮所招致的搏出量减少的现象得到纠正。 但是,既然通过异长调节,搏出量已经恢复原有水平,那么,心室的充盈一不是将同时恢复原水平,异长调节的作用被取消,搏出量不是又因 高动脉压状态的存在而减少了吗?这就是说,搏出量是否会出现时而降低。时而恢复的现象?许多实验都证明,随着搏出题的恢复,心空舒张 末期容积确实也恢复到原水平,但此时,搏出量仍然维持高水平,并不出现再次减少的现象。进一步分析发现,后一情况的出现乃是心肌收缩 能力增强所致,尽管此时产生的张力随负荷的增加而有所增加,但缩短的程度不但可以不降低,反而可能增加。至于这种情况下心肌收缩能力 为什么增强,原因尚不十分清楚,可能是神经机液机制调节的结果。 综上所述,心室后负荷的本身直接着持出量,随后通过异长和等长调节机制,使前负荷和心肌收缩能力与后负荷相匹配,从而使机体得在 在动脉压增高的情况下,能够维持适当的心输出量。这种情况对于机体是有重要生理意义的。但也应看到,此时博出量的维持,是心肌加强收 缩的结果,如果动脉压持续增高,心室肌将因处于收缩强状太而逐渐肥厚,即发生了病理性玫变,随后将导致泵血功能减退。 二)率及其对心输出量的影响 健击成年人的安静状态下,心率平均为每分钟75次(正常范围为每分钟60一100次),不同生理条件下,心率有很大变动,可低到每分钟 400次,高法每合钟200次 心输出量是搏出量与心率的乘积,心率增快,心输出量增加:但这有一定的限度,如果心率增加过快,超过每分钟170“180次,心室充盆 时间明显缩短,率盈量减少,博出题可减少到仅有正常时的一半左右,心确出量亦开始下降。当心率增快但尚未超过此限度时,尽管此时心室 充盈时间有所缩短,但由于回心血量中的绝大部分是在快速充密朗内进入心室的,因此,心室充盘量以及搏出量不致于减少或过分减少,而由 于心庇护增加,每分钟的输出量增加。反之,如心率太慢,低于每分钟40次,心输出量亦减少.这是因为心室舒张蝴过长,心室充盈早已接近 限度,再延长心舒时间也不能相应增加充盈量和博出量.可见,心跳频率最适宜时,心输出量最大,心率过快或过樱,心输出量都会减少。 心率受自主神经的控制,交感神经活动增强时,心率增快:迷走神经活动增强时,心率减慢。影响心率的体液因素主要有循环血液中的肾 上腺和去甲上狼素,以及甲状象素,此外,心率受体温的影响,体温升高1℃,心率将增加12一18次 (白)心避泵功能 、td 分钟75次 能能等广泛通应机体不同生理条件下的代 ,健康成年人静息状态下心率每 左右 时, 搏出量 ad 产是 日使康人心正 贮备 、有相当 出 大出量较正人为 亚左 法3以 力增 而训练有素的运动 为 的3信左右 五.心山音心山音图 心动周期中,心肌收缩、瘦膜启闭、血液加速度和减速度对心血管壁的加压和减压作用以及形成的涡流等因素引起的机械振动,可通过周 围组织传递到胸壁:如将听诊器放在陶壁某些部位,就可以听到声音,称为心音。若用换能器将这些机械振动转换成电信号记录下来,便得到 了心查图, 心音发生在心动周期的某些特定时期,其音调和持续时间也有一定的规律:正常心脏可听到4个心音:即第一、第二,第三和第四心音。多 数情况下只能听到第一和第二心音,在某些健康儿童和青年人也可听到第三心音,40岁以上的使康人也有可能出现第四心音。心胜某些异常活 动可以产生杂音或其它异常心音,因些,听取心音或记录心音图对于心脏疾病的诊浙有一定的意义, 第一心音发生在心缩期,音调低,持续时间相对较长,在心尖搏动处(左第五肋间隙锁骨中线)听得最清范。在心缩期心室射血引起大血 管扩张及产生的涡流发出的低频振动,有及由于房室覆突然关闭所引起的振动,是听诊的第一心音的主要组成成分,因经,通常可用第一心音 人选为心室收缩期开始的标志。笔二心音发生在心正舒张期,颈率较主,持续时间较短。听诊的第二心音主要与主动脉簿的关闭有关,故可用 用标志心室舒张期开始。第三心音发生在快速充盈期末,是一种低频、低振幅的心音。它可能是由于心室快速充空期末,血流充盈减,流速突 然改变,形成一种力使心室壁和要膜发生振动而产生的。第四心音是与心房收缩有关的一组心室收缩期前的振动,故也称心房音。正常心房收 结,听不到声音,但在异常有力的心房收缩和左室壁变硬的情况下,心房收缩使心室充盈的血量增加,心室进一步扩张,引起左室肌及二尖籍 和血液的振动,则可产生第四心音, 第二节心肌的生物电现象和生理特征 心房和心室不停败地进行有顺序的、协调的收缩和舒张交智的活动,是心脏实现泵血功能。推动血液循环的必要条件,而细胞腰的兴奋过 程则是触发收缩反应的始动因素。本节需要阔述的问题是:引起心脏收缩活动的兴奋来自何处?为什么心脏四个腔室能够作协调的收缩活动? 为什么心脏的收缩活动始终是收缩和舒张交替而不出现强直收结?要回答这些问题,必须了解心肌的生理特性,主要是心肌兴奋和兴奋传导的 特征。兴奋和传导是以细的腿的生物电活动为基础的。因此,首先叙述心肌细胞的生物电现象,然后,根据生物电现象分析叙桃心肌兴奋和兴 奋传播的规律和生理意义
例上,可以很容易理解这个问题。当心肌收缩能力不变时,收缩释放的能量是恒定的。如果室内压力升高,表示心肌产生张力消耗的能量相对 增加,用于心肌纤维缩短将相对减少;如果肌肉作等长收缩,释放的能量几乎全部用于产生张力。另一方面,也应该看到,由于动脉压影响了 搏出量,又常常继发地引起一些调节机制的活动。如上述,动脉压的突然升高首先导致搏出量减少,结果造成心室内剩余血量增加,充盈量增 加,后者又再引起异长调节机制的调节,搏出量可以恢复正常水平,即通过异长调节可以使动脉压增高所招致的搏出量减少的现象得到纠正。 但是,既然通过异长调节,搏出量已经恢复原有水平,那么,心室的充盈一不是将同时恢复原水平,异长调节的作用被取消,搏出量不是又因 高动脉压状态的存在而减少了吗?这就是说,搏出量是否会出现时而降低、时而恢复的现象?许多实验都证明,随着搏出题的恢复,心室舒张 末期容积确实也恢复到原水平,但此时,搏出量仍然维持高水平,并不出现再次减少的现象。进一步分析发现,后一情况的出现乃是心肌收缩 能力增强所致。尽管此时产生的张力随负荷的增加而有所增加,但缩短的程度不但可以不降低,反而可能增加。至于这种情况下心肌收缩能力 为什么增强,原因尚不十分清楚,可能是神经机液机制调节的结果。 综上所述,心室后负荷的本身直接着搏出量,随后通过异长和等长调节机制,使前负荷和心肌收缩能力与后负荷相匹配,从而使机体得在 在动脉压增高的情况下,能够维持适当的心输出量。这种情况对于机体是有重要生理意义的。但也应看到,此时搏出量的维持,是心肌加强收 缩的结果,如果动脉压持续增高,心室肌将因处于收缩强状太而逐渐肥厚,即发生了病理性改变,随后将导致泵血功能减退。 (二)心率及其对心输出量的影响 健康成年人的安静状态下,心率平均为每分钟75次(正常范围为每分钟60~100次)。不同生理条件下,心率有很大变动,可低到每分钟 40~50次,高达每分钟200次。 心输出量是搏出量与心率的乘积,心率增快,心输出量增加;但这有一定的限度,如果心率增加过快,超过每分钟170~180次,心室充盆 时间明显缩短,率盈量减少,搏出题可减少到仅有正常时的一半左右,心输出量亦开始下降。当心率增快但尚未超过此限度时,尽管此时心室 充盈时间有所缩短,但由于回心血量中的绝大部分是在快速充盈期内进入心室的,因此,心室充盈量以及搏出量不致于减少或过分减少,而由 于心庇护增加,每分钟的输出量增加。反之,如心率太慢,低于每分钟40次,心输出量亦减少。这是因为心室舒张期过长,心室充盈早已接近 限度,再延长心舒时间也不能相应增加充盈量和搏出量。可见,心跳频率最适宜时,心输出量最大,心率过快或过慢,心输出量都会减少。 心率受自主神经的控制,交感神经活动增强时,心率增快;迷走神经活动增强时,心率减慢。影响心率的体液因素主要有循环血液中的肾 上腺和去甲肾上腺素,以及甲状腺素。此外,心率受体温的影响,体温升高1℃,心率将增加12~18次。 (三)心脏泵功能的贮备 心脏的泵血功能能够广泛适应机体不同生理条件下的代谢需要,表现为心输出量可随机体代谢增长而增加。健康成年人静息状态下心率每 分钟75次,搏出量约70ml,心输出量为5L左右。强体力劳动时,心率可达每分钟180~200次,搏出量可增加到150ml左右,心输出量可达25~ 30L,为静息时的5~6倍。心脏每分钟能射出的最大血量,称最大输出量。它反映心脏的健康程度。由上可以看出,在平时,心输出量产是最 大的,但能够在需要时成倍地增长,表明健康人心脏泵血功能有一定的贮备力量。心输出量随机体代谢需要而增加的能力,称为泵功能贮备, 或心力贮备。健康人有相当大的心力贮备,而某些心脏疾患的病人,静息时心输出量与健康人没明显差别,尚能够满足静息状态下代谢的需 要,但在代谢活动增强时,输出量却不以相应增加,最大输出量较正常人为低;而训练有素的运动员,心脏的最大输出量远比一般人为主,可 达35L以上,为静息时的8倍左右。 五、心音心音图 心动周期中,心肌收缩、瓣膜启闭、血液加速度和减速度对心血管壁的加压和减压作用以及形成的涡流等因素引起的机械振动,可通过周 围组织传递到胸壁;如将听诊器放在胸壁某些部位,就可以听到声音,称为心音。若用换能器将这些机械振动转换成电信号记录下来,便得到 了心音图。 心音发生在心动周期的某些特定时期,其音调和持续时间也有一定的规律;正常心脏可听到4个心音:即第一、第二、第三和第四心音。多 数情况下只能听到第一和第二心音,在某些健康儿童和青年人也可听到第三心音,40岁以上的健康人也有可能出现第四心音。心脏某些异常活 动可以产生杂音或其它异常心音。因些,听取心音或记录心音图对于心脏疾病的诊断有一定的意义。 第一心音发生在心缩期,音调低,持续时间相对较长,在心尖搏动处(左第五肋间隙锁骨中线)听得最清楚。在心缩期心室射血引起大血 管扩张及产生的涡流发出的低频振动,有及由于房室瓣突然关闭所引起的振动,是听诊的第一心音的主要组成成分,因经,通常可用第一心音 人选为心室收缩期开始的标志。第二心音发生在心脏舒张期,频率较主,持续时间较短。听诊的第二心音主要与主动脉瓣的关闭有关,故可用 用标志心室舒张期开始。第三心音发生在快速充盈期末,是一种低频、低振幅的心音。它可能是由于心室快速充盈期末,血流充盈减,流速突 然改变,形成一种力使心室壁和瓣膜发生振动而产生的。第四心音是与心房收缩有关的一组心室收缩期前的振动,故也称心房音。正常心房收 缩,听不到声音,但在异常有力的心房收缩和左室壁变硬的情况下,心房收缩使心室充盈的血量增加,心室进一步扩张,引起左室肌及二尖瓣 和血液的振动,则可产生第四心音。 第二节 心肌的生物电现象和生理特征 心房和心室不停歇地进行有顺序的、协调的收缩和舒张交替的活动,是心脏实现泵血功能、推动血液循环的必要条件,而细胞膜的兴奋过 程则是触发收缩反应的始动因素。本节需要阐述的问题是:引起心脏收缩活动的兴奋来自何处?为什么心脏四个腔室能够作协调的收缩活动? 为什么心脏的收缩活动始终是收缩和舒张交替而不出现强直收缩?要回答这些问题,必须了解心肌的生理特性,主要是心肌兴奋和兴奋传导的 特征。兴奋和传导是以细胞膜的生物电活动为基础的。因此,首先叙述心肌细胞的生物电现象,然后,根据生物电现象分析叙述心肌兴奋和兴 奋传播的规律和生理意义

心肌细胞的类型组成心脏的心肌细胞并不是同一类型的,根据它们的组织学特点、电生理特性以及功能上的区别,粗略地分为两大类型 两类心肌细跑分别实现一定的职能,互相配合,完成心脏的整体活动。一类是普通的心肌细抱,包括心房肌和心室肌,含有丰富的肌原纤维 执行收缩功能,故又称为工作细跑。工作细跑不能自动地产生节律性兴奋,即不具有自动节律性:但它具有兴奋性,可以在外来刺激作用下行 生兴奋:也具有传导兴奋的能力,但是,与相应的特殊传导组织作比较,传导性较低。另一类是一些特殊分化了的心肌细,组成心脏的特殊 传导系统:其中主要包括细孢和哺肯野细胞,它们除了具有兴奋性和传导性之外,还具有自动产生节律性兴奋的能力,故称为自律细跑,它 们含肌原纤维甚小或完全缺乏,故收缩功能已基本丧失,还有一种细孢位于特殊传导系统的结区,既不具有收缩功能,也没有自律性,只保留 了很低的传导性,是传导系统中的非自律细胞,特殊传导系统是心脏内发生兴奋和传播兴奋的组织,起着控制心脏节律性活动的作用。 心脏特殊传导系统的组成和分布心脏的特殊传导系统由不同类型的特殊分化的心肌细胞所组成。包括窦房结、房室交界、房室束和未梢浦 肯野纤维网(图45)· 窦房结:位于右心房和上腔静脉连接处,主要含有P细胞和过演细胞。P细孢是自律细孢,位于安房结中心部分:过渡细泡位于周边部分 不具有自律性,其作用是将P细跑自动产生的兴奋向外传播到心房肌, 房室交界:又称为房室结区,是心房与心室之间的特殊传导组织,是心房兴奋传入心室的通道。房室交界主要包括以下三个功能区域: 房结区:位于心房和结区之间,具有传导性和自律性。 结区:相当于光学显微镜所见的房室结,具有传导性,无自律性, 结希区:位于结区和希氏束之间,只有传导性和自律性」 房室束(仅称希氏束)及其分支:房室束走行于室间隔内,在室间阴膜部开始分为左右两支,右束支较细,沿途分支少,分布于右心室 左束支显带状,分支多,分布于左心室,房室束主要含浦肯野细泡. 1.0 000 时闻(ms) 图45心蓝各部分心肌细跑的跨模电位和兴奋传导速度 SAM:实房结AM:心房肌AVN;结区BH:希氏束PE;哺情野纤维 TPF:末梢浦惰野纤维VM:心室肌传导速度单位m5 浦肯野纤维网:是左右束支的最后分支,由于分支很多,形成网状,密布于左右心室的心内膜下,并垂直向心外膜侧伸延,再与普通心室 肌细胞相连接。房室束及未梢浦肯野纤维网的作用,是将心房传来的兴奋迅速传播到整个心室。 关于是否存在心传导的问题。争论很多。60年代 ac提出在窦房结和房室结区之间有三条由浦肯野细胞构成的心房传导束,分别 整齐,因此其传导速度较其它部位心房肌(这些心房肌被右心房壁上腔静脉开口卵圆窝所形成的孔穴所分制,形成新续状)为快,从而在功能 上构成了将房结兴奋快速传播到房室交界处的所调优均传导通路D心心 ial pathy 一、心肌细胞的生物电现象 与骨肌相此,心肌细跑的跨膜电位在波形上和形成机制上要复杂得多:不但如此,上述不同类型的心肌细的骑膜电位(45),不仅 幅度和持续时 间各不相 而且波形和形成的商子基础也有一定的差别:各类心肌细胞电活动的不一致性,是心脏兴奋的产生以及兴奋向整个 心旺传播过程中表现出特殊规律的原因 (一)工作细胞的跨膜电位及其形成机制 1.静息电位和动作电位人和哺乳动物的心室肌细胞和骨略肌细胞一样,在静息状态下膜两侧呈极化状态,膜内电位比膜外电位约低 90mV,但两者的动作电位有明显不同.。骨略肌细跑动作电位的时程很短,仅持续几个毫秒,复极速度与去极速度几乎相等,记录曲线呈升支 和降支基本对称的尖锋状。心空肌细胞动作电位的主要特征在于复极过程比较复杂,持续时间很长,动作电位降支与升支很不对称。通常用 0、1、2,3、4等数字分别代表心室肌细跑动作电位和静息电位的各个时期。 (1)除极(去极)过程:除极过程又称0期。在适宜的外来刺激作用下,心室肌细胞发生兴奋,膜内电位由静息状态下的-90mV迅速上升 到+30mV左右,即肌膜两原有的极化状态被消除井呈极化倒转,构成动作电位的升支,除极相很短暂,仅占1-2m5,而且除极幅度很大,为 120mV:可见,心室肌细胞的除极速度很快,膜电位的最大变化速率可达300-1000Vs
心肌细胞的类型组成心脏的心肌细胞并不是同一类型的,根据它们的组织学特点、电生理特性以及功能上的区别,粗略地分为两大类型: 两类心肌细胞分别实现一定的职能,互相配合,完成心脏的整体活动。一类是普通的心肌细胞,包括心房肌和心室肌,含有丰富的肌原纤维, 执行收缩功能,故又称为工作细胞。工作细胞不能自动地产生节律性兴奋,即不具有自动节律性;但它具有兴奋性,可以在外来刺激作用下产 生兴奋;也具有传导兴奋的能力,但是,与相应的特殊传导组织作比较,传导性较低。另一类是一些特殊分化了的心肌细胞,组成心脏的特殊 传导系统;其中主要包括P细胞和哺肯野细胞,它们除了具有兴奋性和传导性之外,还具有自动产生节律性兴奋的能力,故称为自律细胞,它 们含肌原纤维甚小或完全缺乏,故收缩功能已基本丧失。还有一种细胞位于特殊传导系统的结区,既不具有收缩功能,也没有自律性,只保留 了很低的传导性,是传导系统中的非自律细胞,特殊传导系统是心脏内发生兴奋和传播兴奋的组织,起着控制心脏节律性活动的作用。 心脏特殊传导系统的组成和分布心脏的特殊传导系统由不同类型的特殊分化的心肌细胞所组成。包括窦房结、房室交界、房室束和末梢浦 肯野纤维网(图4-5)。 窦房结:位于右心房和上腔静脉连接处,主要含有P细胞和过渡细胞。P细胞是自律细胞,位于窦房结中心部分;过渡细胞位于周边部分, 不具有自律性,其作用是将P细胞自动产生的兴奋向外传播到心房肌。 房室交界:又称为房室结区,是心房与心室之间的特殊传导组织,是心房兴奋传入心室的通道。房室交界主要包括以下三个功能区域: 房结区:位于心房和结区之间,具有传导性和自律性。 结区:相当于光学显微镜所见的房室结,具有传导性,无自律性。 结希区:位于结区和希氏束之间,具有传导性和自律性。 房室束(又称希氏束)及其分支:房室束走行于室间隔内,在室间隔膜部开始分为左右两支,右束支较细,沿途分支少,分布于右心室, 左束支呈带状,分支多,分布于左心室,房室束主要含浦肯野细胞。 图4-5 心脏各部分心肌细胞的跨膜电位和兴奋传导速度 SAM:窦房结 AM:心房肌 AVN;结区 BH:希氏束 PE;哺肯野纤维 TPF:末梢浦肯野纤维 VM:心室肌传导速度单位m/s 浦肯野纤维网:是左右束支的最后分支,由于分支很多,形成网状,密布于左右心室的心内膜下,并垂直向心外膜侧伸延,再与普通心室 肌细胞相连接。房室束及末梢浦肯野纤维网的作用,是将心房传来的兴奋迅速传播到整个心室。 关于是否存在心房传导束的问题,争论很多。60年代,Janes提出在窦房结和房室结区之间有三条由浦肯野细胞构成的心房传导束,分别称 前、中、后结间束,其兴奋传导速度比一般心房肌为快。但是,近20年来的研究未能证实心房内有形态结构上不同于心房(工作)细胞的特殊 传导组织组成的心房传导束存在;另一方面,研究结果表明,在右心房的某些部位(如卵圆窝前方和界嵴处)心房肌纤维排列方向一致,结构 整齐,因此其传导速度较其它部位心房肌(这些心房肌被右心房壁上腔静脉开口卵圆窝所形成的孔穴所分割,形成断续状)为快,从而在功能 上构成了将窦房结兴奋快速传播到房室交界处的所谓优势传导通路(preferential pathway) 。 一、心肌细胞的生物电现象 与骨骼肌相比,心肌细胞的跨膜电位在波形上和形成机制上要复杂得多;不但如此,上述不同类型的心肌细胞的跨膜电位(图4-5),不仅 幅度和持续时间各不相同,而且波形和形成的离子基础也有一定的差别;各类心肌细胞电活动的不一致性,是心脏兴奋的产生以及兴奋向整个 心脏传播过程中表现出特殊规律的原因。 (一)工作细胞的跨膜电位及其形成机制 1.静息电位和动作电位人和哺乳动物的心室肌细胞和骨骼肌细胞一样,在静息状态下膜两侧呈极化状态,膜内电位比膜外电位约低 90mV,但两者的动作电位有明显不同。骨骼肌细胞动作电位的时程很短,仅持续几个毫秒,复极速度与去极速度几乎相等,记录曲线呈升支 和降支基本对称的尖锋状。心室肌细胞动作电位的主要特征在于复极过程比较复杂,持续时间很长,动作电位降支与升支很不对称。通常用 0、1、2、3、4等数字分别代表心室肌细胞动作电位和静息电位的各个时期。 (1)除极(去极)过程:除极过程又称0期。在适宜的外来刺激作用下,心室肌细胞发生兴奋,膜内电位由静息状态下的-90mV迅速上升 到+30mV左右,即肌膜两侧原有的极化状态被消除并呈极化倒转,构成动作电位的升支。除极相很短暂,仅占1-2ms,而且除极幅度很大,为 120mV;可见,心室肌细胞的除极速度很快,膜电位的最大变化速率可达800-1000V/s

(2)复极过程:当心室细胞除极达到顶峰之后,立即开始复极,但整个复极过程比较缓慢,包括电位变化曲线的形态和形成机制均不相同 的三个阶段: 1期复极:在复极初期,仅出现部分复极,膜内电位由+30mV迅速下降到0mV左右,故1期又称为快速复极初期,占时约10ms,0期除极和 1期复极这两个时期的膜电位的变化速度都很快,记录图形上表现为尖锋状,故在心肌细胞习惯上常把这两部分合称为锋电位。 2期复极:当1期复极膜内电位达到0mV左右之后,复极过程就变得非常缓慢,膜内电位基本上停滞于0mV左右,细胞膜两侧显等电位状 态,记录图形比较平坦,故复极2期又称为坪或平台期,持续约100-150m5,是整个动作电位持续时间长的主要原因,是心室肌细跑以及其它心 肌细胞的动作电位区别于骨略肌和神经纤维的主要特征。 3期复极:2期复极过程中,随着时间的进展,膜内电位以较慢的速度由0mV逐渐下降,延续为3期复极,2期和3期之间没有明显的界限。 在3期,细跑摸复极速度加快,膜内电位由0mV左右较快地下降到-90mV,完成复极化过程,故3期又称为快速复极未期,占时约100-150ms, 4期:4期是膜复极完毕、膜电位恢复后的时期。在心室肌细胞或其它非自律细胞,4期内膜电位稳定于静息电位水平,因此,4期又可称为 静息期 2。形成机制与骨酪肌一样,离子在细孢膜两侧不均匀分布所形成的浓度梯度(浓度差)(表41),驱动相应离子经过当时开放的细胞膜 上特殊离子通道的跨膜扩散,是心肌细胞跨膜电位形成的主要基础,只是由于心肌细胞膜上具有数目较多的高子通道,跨膜电位形成机制中涉 及的离子流远比骨胳肌要复杂得多。在电生理学中,电流的方向以正离子在膜两侧的流动方向来命名,正离子外流或负离子内流称外向电流, 离子内流或负离子外流称内向电流。外向电流导致膜内电位向负电性转化,促使膜复极,内向电流导致膜内电位向正电性转化,促使膜除 极 表41心肌细胞中各种主要离子的浓度及平衡电位值 子 内外比值 : 10 135 3 除离子跨膜扩散之外,由细抱上离子泵所实现的离子主动转运和离子交换,在心肌细胞电活动中也占有重要地位, 心室肌细泡静息电位的形成机制与骨路肌相同,也就是说,尽管肌膜两侧上述几种离子都存在有浓度梯度,但静息状态下肌膜对水*的通透 性较高,而对其它离子的通透性很低,因此,K+顺其浓度梯度由膜内向膜外扩散所达到的平衡电位,是静息电位的主要来源。 肌膜钠通道的大量开放和膜两侧浓度梯度及电位梯度的动从而出现、a快速内流,是心室肌细0胡去极形成的原因。进一步对整个去极 过程进行分析就可以看到,与骨酪肌一样,在外来刺激作用下 ,首先引起部分电压门控式Na通道开放和少量N内流,造成肌要部分去极化 谟电位绝对值下隆:而当膜电位由静息水平(膜内-90mV)去极化到国电位水平(膜内-70mV)时,膜上Na通道开放概率明显增加,出现开 生性N妇内流(参看第二章),于是N顺其浓度梯度和电位梯度由要外快速进入膜内,进一步使膜去极化,膜内电位向正电性转化。决定0期 去极的Na通道是一种快通道,它不但激活、开放的速度很快,而且激活后很快就失活,当膜除极到一定程度(oV左右)时,N通道就开 始失活而关闭,最后终止Na的继续内流。快Na+通道可被河豚毒(TTX)所阻断。由于Na通道激活速度非常之快,又有再生性循环出现,这 就是心室肌细胞0期去极速度很快,动作电位升支非常陡峭的原因。正因为如此,从电生理特性上,尤其是根据0期除极的速率,将心室肌细胞 (以及具有同样特征的心肌细胞)称为快反应细胞,其动作电位称为快反应电位,以区别于以后将要介绍的慢反应细胞和慢反应电位。 复极1期是在0期除极之后出现的快速而短暂的复极期,此时快钠通道已经失活,同时激活一种一过性外向电流(),从而使膜迅速复极 到平台期电位水平(0-20mV)至于1o的离子成分,70年代曾认为是Cr(即Cr内流)近年来,根据0可被四乙基铵和4-氨基此啶 K*通道阳滞剂所阻断的研究资料,认为K*才是的主要离子成分。也就是说,由K负载的一过性外向电流是动作电位初期快速复极的主要原 因。目前对0的通道特征尚不十分清楚,但有资料提示,膜除极和细胞内C2+都可以使0的通道激活 平台期初期,膜电位稳定于OmV左右, 两种电流处于相对平衡状态,随后,内向电流逐新减弱,分 电流 ,总和的结果是出现 流,导致膜电位缓慢地向膜内负电性转化。电压钳研究结果表明,在心室肌等快反应细胞,平台期外向离子流是由K*携带的(称1)·静息 状态下,K*通道的通透性很高,在0明除极过程中,K*的通透性显著下降,K*外流大大减少,除极相结束时,K的通透性并不是立即恢复到 静息状态下的那种高水平,而是极其缓慢地、部分地恢复,K*外流也就由初期的低水平而慢慢增加(图46),平台期内向离子流主要是由 C✉2+(以及N负载的。已经证明,心肌细胞摸上有一种电压门控式的慢Ca2通道,当膜除极到40mV时被激活,Ca2顺其浓度梯度向膜内缓 慢扩散从而倾向于使膜除极,与此同时,上述微弱的K*外流倾向于使膜复极化,在平台期早期,C:+的内流和K*的外流所负裁的跨膜正电荷 时相等,膜电位稳定于1期复极所达到的电位水平。随着时间推移,C2“通道逐渐失活,K*外流逐渐增加,其结果,出膜的净正电荷量逐渐增 加,膜内电位于是逐渐下降,形成平台期晚期。此后,C2+通道完全失活,内向离子流终止,外向K*流进一步增强。平台蝴延续为复极3期 电位较快地回到静息水平,完成复极化过程
(2)复极过程:当心室细胞除极达到顶峰之后,立即开始复极,但整个复极过程比较缓慢,包括电位变化曲线的形态和形成机制均不相同 的三个阶段: 1期复极:在复极初期,仅出现部分复极,膜内电位由+30mV迅速下降到0mV左右,故1期又称为快速复极初期,占时约10ms。0期除极和 1期复极这两个时期的膜电位的变化速度都很快,记录图形上表现为尖锋状,故在心肌细胞习惯上常把这两部分合称为锋电位。 2期复极:当1期复极膜内电位达到0mV左右之后,复极过程就变得非常缓慢,膜内电位基本上停滞于0mV左右,细胞膜两侧呈等电位状 态,记录图形比较平坦,故复极2期又称为坪或平台期,持续约100-150ms,是整个动作电位持续时间长的主要原因,是心室肌细胞以及其它心 肌细胞的动作电位区别于骨骼肌和神经纤维的主要特征。 3期复极;2期复极过程中,随着时间的进展,膜内电位以较慢的速度由0mV逐渐下降,延续为3期复极,2期和3期之间没有明显的界限。 在3期,细胞膜复极速度加快,膜内电位由0mV左右较快地下降到-90mV,完成复极化过程,故3期又称为快速复极末期,占时约100-150ms 。 4期:4期是膜复极完毕、膜电位恢复后的时期。在心室肌细胞或其它非自律细胞,4期内膜电位稳定于静息电位水平,因此,4期又可称为 静息期。 2.形成机制与骨骼肌一样,离子在细胞膜两侧不均匀分布所形成的浓度梯度(浓度差)(表4-1)、驱动相应离子经过当时开放的细胞膜 上特殊离子通道的跨膜扩散,是心肌细胞跨膜电位形成的主要基础,只是由于心肌细胞膜上具有数目较多的离子通道,跨膜电位形成机制中涉 及的离子流远比骨骼肌要复杂得多。在电生理学中,电流的方向以正离子在膜两侧的流动方向来命名,正离子外流或负离子内流称外向电流, 正离子内流或负离子外流称内向电流。外向电流导致膜内电位向负电性转化,促使膜复极,内向电流导致膜内电位向正电性转化,促使膜除 极。 表4-1 心肌细胞中各种主要离子的浓度及平衡电位值 离 子 浓度(mmo1/L) 细胞内液 细胞外液 内/外比值 平衡电位(mV) (由Nernst公式计算) Na+ 30 140 1:4,6 +41 K + 140 4 35:1 -94 Ca2+ 10-4 2 1:20,000 +132 CI- 30 104 1:3.5 -33 除离子跨膜扩散之外,由细胞上离子泵所实现的离子主动转运和离子交换,在心肌细胞电活动中也占有重要地位。 心室肌细胞静息电位的形成机制与骨骼肌相同,也就是说,尽管肌膜两侧上述几种离子都存在有浓度梯度,但静息状态下肌膜对K +的通透 性较高,而对其它离子的通透性很低,因此,K +顺其浓度梯度由膜内向膜外扩散所达到的平衡电位,是静息电位的主要来源。 肌膜钠通道的大量开放和膜两侧浓度梯度及电位梯度的驱动从而出现Na+快速内流,是心室肌细胞0期去极形成的原因。进一步对整个去极 过程进行分析就可以看到,与骨骼肌一样,在外来刺激作用下,首先引起部分电压门控式Na+通道开放和少量Na+内流,造成肌膜部分去极化, 膜电位绝对值下降;而当膜电位由静息水平(膜内-90mV)去极化到阈电位水平(膜内-70mV)时,膜上Na+通道开放概率明显增加,出现再 生性Na+内流(参看第二章),于是Na+顺其浓度梯度和电位梯度由膜外快速进入膜内,进一步使膜去极化,膜内电位向正电性转化。决定0期 去极的Na+通道是一种快通道,它不但激活、开放的速度很快,而且激活后很快就失活,当膜除极到一定程度(omV左右)时,Na+通道就开 始失活而关闭,最后终止Na+的继续内流。快Na+通道可被河豚毒(TTX)所阻断。由于Na+通道激活速度非常之快,又有再生性循环出现,这 就是心室肌细胞0期去极速度很快,动作电位升支非常陡峭的原因。正因为如此,从电生理特性上,尤其是根据0期除极的速率,将心室肌细胞 (以及具有同样特征的心肌细胞)称为快反应细胞,其动作电位称为快反应电位,以区别于以后将要介绍的慢反应细胞和慢反应电位。 复极1期是在0期除极之后出现的快速而短暂的复极期,此时快钠通道已经失活,同时激活一种一过性外向电流(Ito),从而使膜迅速复极 到平台期电位水平(0~-20mV)。至于Ito的离子成分,70年代曾认为是Cl -(即Cl -内流)。近年来,根据Ito可被四乙基铵和4-氨基吡啶等 K +通道阻滞剂所阻断的研究资料,认为K +才是Ito的主要离子成分。也就是说,由K +负载的一过性外向电流是动作电位初期快速复极的主要原 因。目前对Ito的通道特征尚不十分清楚,但有资料提示,膜除极和细胞内Ca 2+都可以使Ito的通道激活。 平台期初期,膜电位稳定于0mV左右,随后才非常缓慢地复极。膜电位的这种特征是由于平台期同时有内向电流和外向电流存在,初期, 两种电流处于相对平衡状态,随后,内向电流逐渐减弱,外向电流逐渐增强,总和的结果是出现一种随时间推移而逐渐增强的、微弱的外向电 流,导致膜电位缓慢地向膜内负电性转化。电压钳研究结果表明,在心室肌等快反应细胞,平台期外向离子流是由K +携带的(称Ik1)。静息 状态下,K +通道的通透性很高,在0期除极过程中,K +的通透性显著下降,K +外流大大减少,除极相结束时,K +的通透性并不是立即恢复到 静息状态下的那种高水平,而是极其缓慢地、部分地恢复,K +外流也就由初期的低水平而慢慢增加(图4-6)。平台期内向离子流主要是由 Ca 2+ (以及Na+ )负载的。已经证明,心肌细胞膜上有一种电压门控式的慢Ca 2+通道,当膜除极到-40mV时被激活,Ca 2+顺其浓度梯度向膜内缓 慢扩散从而倾向于使膜除极,与此同时,上述微弱的K +外流倾向于使膜复极化,在平台期早期,Ca 2+的内流和K +的外流所负载的跨膜正电荷 时相等,膜电位稳定于1期复极所达到的电位水平。随着时间推移,Ca 2+通道逐渐失活,K +外流逐渐增加,其结果,出膜的净正电荷量逐渐增 加,膜内电位于是逐渐下降,形成平台期晚期。此后,Ca 2+通道完全失活,内向离子流终止,外向K +流进一步增强,平台期延续为复极3期, 膜电位较快地回到静息水平,完成复极化过程

Na 图46心室肌细胞跨膜电位及其形成的离子机制 RMP:静息要电位P:电位 肌膜上有C✉+遥道,是心室肌细胞和其它心肌细胞的重要特征。大量研究表明:①从一个心肌细跑的总体而言(不是从单个通道而言) C:+通道的激活、失活。以及再复活所需时间均比八、a通道要长,经C:+通道跨模的C:+内流。起始慢,平均持续时间也较长,因此相应称为 慢通道和慢内向离子流②慢通道也是电压门控式的,激活慢通道的闲电位水平(5035mV)高于快N通道70~-55mV):它对某些 理化因素的感性和反应性不于快语道。可被M“和多 Ca24阳断剂(如异博定 D600等)所阳齿 ,而对于可以阻断快通道的河毒和年 膜的持续低极化状态(膜内电位50M左右)却并不敏感,各种心肌细胞的肌膜上都具有这种慢通道,由此形成的跨膜离子流,是决定心肌 细胞电活动以及心室肌等快反应细胞动作电位平台期的最重要的内向离子流之一, 平台期之后,膜的复极逐渐加速,因此时C2+通道已经失活,在平台期已经激活的外向K*流出现随时间而递增的趋势。其原因是,3期的 一复授程再生任的,水的金透是速叠内电位向负电性转化。而展内电位腾负,《外花亮送帽高。这种正反情过程号孩要的复技道来 在4期内,心室肌细抱膜电位基本上稳定于静息电位水平,但是,离子的跨膜转运仍然在活跃进行。因为,动作电位期问有Na'和C+进入 细胞内,而K*外流出细跑,因此,只有从细胞内排出多余的、和C2,并摄入K*才能恢复细胞内外离子的正常浓度梯度,保持心肌细胞的正 常兴奋性。这种离子转运是逆着浓度梯度进行的主动转运过程。像骨略肌一样,通过肌膜上N'K*泵的作用,将N的外运和K+的内运互相 联形成NaK转运,同时实现Na和K的主动转运。关于主动转运C的转运机制,还没有完全弄清楚。目前大多数作者认为,C2的逆浓 度梯度的外运是与Na的顺浓度的内流相合进行的。形成Na.Ca2+交换。Ca2的这种主动转运是由Na的内向性浓度梯度提供能量的。由于 Na*内向性浓度悌度的维持是依靠NaK泵而实现的,因此,Ca2主动转运也是由NaK泵提供能量的。在4期开始后,膜的上述主动转运 加强,细跑内外离子浓度梯度得以恢复。总的来看,这时转运过程引起的跨膜交换的电荷量基本相等,因此,膜电位不受影响而能维持稳 定 (仁)自律细胞的跨膜电位及其形成机制 在没有外来刺激时,工作细胞不能产生动作电位,在外来刺激作用下,产生一次动作电位,但两次动作电位之间膜电位是稳定不变的。而 在自律细跑,当动作电位3期复极未期达到最大值(称最大复极电位)之后,4期的膜电位并不稳定于这一水平,而是立即开始自动除极,除极 达司电位后引起兴奋,出现另一个动作电位。这种现象,周而复始,动作电位就不断地产生。出现于4期的这种自动除极过程,具有随时间而 递增的特点,其除极速度远较蝴除极缓慢;不同类型的自律细胞4期除极速度参差不一,但同类自律细孢4期除极速度比较恒定。这种4蝴自动 除极(亦称4期缓慢除极或缓慢舒张期除极】,是自律细胞产生自动节律性兴奋的基础 根据细胞膜除极的跨膜电流的基本规律可分析自律细胞4期自动除极形成的机制。不难推测,自律细胞由于净外向电流使膜复极(3期)达 最大复极电位后,在4期中又出现一种逐渐增强的净内向电流,从而使膜内正电位逐渐增加,膜便逐渐除极。这种进行性净内向电流的产生, 有以下三种可能的原因:@内向电流的逐渐增强;②外向电流的逐渐衰退:®两者兼有。不同类型的自律细跑,4期自动除极都是由这种进行 性净内向电流所引起,但构成净内向电流的离子流的方向和离子本质并不完全相同。 1,浦情野细胞浦肯野细跑是一种快反应自律细胞。作为一种快反应型细胞,它的动作电位的形态与心室肌细胞相似,产生的离子基础也 本相同 关于浦情野细 胞4期自动除极 成的机制,80年代研究资料表明,在浦 野细胞,陆着复极的进行,导致膜复极的外向、 一一在灵电可卫来一片时日花移而逐布棉强的内电速的图刊·通道在动作电位限复吸电达:心生右开德。 开放,其激活程度随着复极的进行、膜内负电性的增加而增加,至·100mV左右就充分激活。因此,内向电流表现出时间依从性增强,膜的除 极程度因而也随时间而增加,一旦达到同电位水平,使又产生另一次动作电位,与此同时,这种内向电流在膜除极达50mV左右因通道失活而 中止,可见,动作电位的复极期膜电位本身是引起这种内向电流启动和发展的因素,内向电流的产生和增强导致膜的进行性除极,而要的除极
图4-6 心室肌细胞跨膜电位及其形成的离子机制 RMP:静息膜电位 TP :阈电位 肌膜上有Ca 2+通道,是心室肌细胞和其它心肌细胞的重要特征。大量研究表明:①从一个心肌细胞的总体而言(不是从单个通道而言), Ca 2+通道的激活、失活,以及再复活所需时间均比Na通道要长,经 Ca 2+通道跨膜的Ca 2+内流,起始慢,平均持续时间也较长。因此相应称为 慢通道和慢内向离子流;②慢通道也是电压门控式的,激活慢通道的阈电位水平(-50~-35mV)高于快Na通道(-70~-55mV);③它对某些 理化因素的敏感性和反应性不同于快通道,可被Mn 2+和多种Ca 2+阻断剂(如异博定,D-600等)所阻断,而对于可以阻断快通道的河豚毒和细 胞膜的持续低极化状态(膜内电位-50Mv左右)却并不敏感。各种心肌细胞的肌膜上都具有这种慢通道,由此形成的跨膜离子流,是决定心肌 细胞电活动以及心室肌等快反应细胞动作电位平台期的最重要的内向离子流之一。 平台期之后,膜的复极逐渐加速,因此时Ca 2+通道已经失活,在平台期已经激活的外向K +流出现随时间而递增的趋势。其原因是,3期的 复极K +流是再生性的,K+的外流促使膜内电位向负电性转化,而膜内电位越负,K+外流就越增高。这种正反馈过程,导致膜的复极越来越 快,直至复极化完成。 在4期内,心室肌细胞膜电位基本上稳定于静息电位水平,但是,离子的跨膜转运仍然在活跃进行。因为,动作电位期间有Na+和Ca 2+进入 细胞内,而K +外流出细胞,因此,只有从细胞内排出多余的Na+和Ca 2+,并摄入K +才能恢复细胞内外离子的正常浓度梯度,保持心肌细胞的正 常兴奋性。这种离子转运是逆着浓度梯度进行的主动转运过程。像骨骼肌一样,通过肌膜上Na+ -K +泵的作用,将Na+的外运和K+的内运互相耦 联形成Na+ -K +转运,同时实现Na+和K +的主动转运。关于主动转运Ca 2+的转运机制,还没有完全弄清楚。目前大多数作者认为,Ca 2+的逆浓 度梯度的外运是与Na+的顺浓度的内流相耦合进行的。形成Na+ -Ca 2+交换。Ca 2+的这种主动转运是由Na+ 的内向性浓度梯度提供能量的,由于 Na+内向性浓度梯度的维持是依靠Na+ -K +泵而实现的,因此,Ca 2+主动转运也是由Na+ -K +泵提供能量的。在4期开始后,膜的上述主动转运功 能加强,细胞内外离子浓度梯度得以恢复。总的来看,这时转运过程引起的跨膜交换的电荷量基本相等,因此,膜电位不受影响而能维持稳 定。 (二)自律细胞的跨膜电位及其形成机制 在没有外来刺激时,工作细胞不能产生动作电位,在外来刺激作用下,产生一次动作电位,但两次动作电位之间膜电位是稳定不变的。而 在自律细胞,当动作电位3期复极未期达到最大值(称最大复极电位)之后,4期的膜电位并不稳定于这一水平,而是立即开始自动除极,除极 达阈电位后引起兴奋,出现另一个动作电位。这种现象,周而复始,动作电位就不断地产生。出现于4期的这种自动除极过程,具有随时间而 递增的特点,其除极速度远较0期除极缓慢;不同类型的自律细胞4期除极速度参差不一,但同类自律细胞4期除极速度比较恒定。这种4期自动 除极(亦称4期缓慢除极或缓慢舒张期除极),是自律细胞产生自动节律性兴奋的基础。 根据细胞膜除极的跨膜电流的基本规律可分析自律细胞4期自动除极形成的机制。不难推测,自律细胞由于净外向电流使膜复极(3期)达 最大复极电位后,在4期中又出现一种逐渐增强的净内向电流,从而使膜内正电位逐渐增加,膜便逐渐除极。这种进行性净内向电流的产生, 有以下三种可能的原因:①内向电流的逐渐增强;②外向电流的逐渐衰退;③两者兼有。不同类型的自律细胞,4期自动除极都是由这种进行 性净内向电流所引起,但构成净内向电流的离子流的方向和离子本质并不完全相同。 1.浦肯野细胞浦肯野细胞是一种快反应自律细胞。作为一种快反应型细胞,它的动作电位的形态与心室肌细胞相似,产生的离子基础也基 本相同。 关于浦肯野细胞4期自动除极形成的机制,80年代研究资料表明,在浦肯野细胞,随着复极的进行,导致膜复极的外向K +电流逐渐衰减, 而同时在膜电位4期可记录到一种随时间推移而逐渐增强的内向电流(I f)(图4-7)。I f通道在动作电位3期复极电位达-60mV左右开始被激活 开放,其激活程度随着复极的进行、膜内负电性的增加而增加,至-100mV左右就充分激活。因此,内向电流表现出时间依从性增强,膜的除 极程度因而也随时间而增加,一旦达到阈电位水平,便又产生另一次动作电位,与此同时,这种内向电流在膜除极达-50mV左右因通道失活而 中止。可见,动作电位的复极期膜电位本身是引起这种内向电流启动和发展的因素,内向电流的产生和增强导致膜的进行性除极,而膜的除极
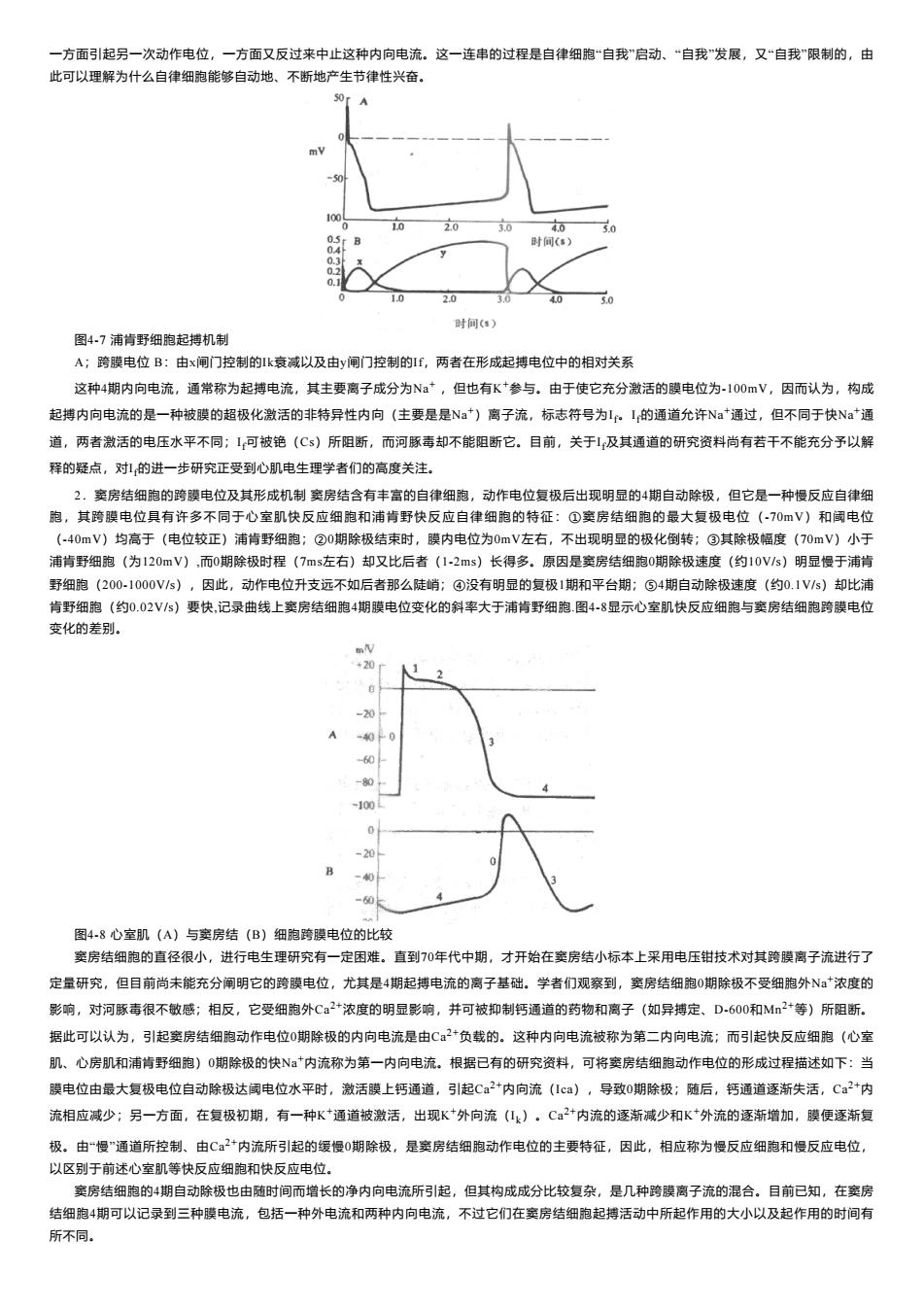
一方面引起另一次动作电位, 一方面又反过来中止这种内向电流。这一连串的过程是自律细胞“自我”启动、“自我”发展,又“自我“限制的,由 此可以理解为什么自律细胞能够自动地、不断地产生节律性兴奋。 A 时阿(a) 图47浦情野细陶起持机制 A:跨膜电位B:由x闸门控制的k衰减以及由y闸门控制的,两者在形成起电位中的相对关系 这种4蝴内向电流。通常称为起搏电流,其主要离子成分为Na,但也有K+参与,由于使它充分激活的膜电位为-10V,因而认为,构成 起搏内向电流的是一种被膜的超极化激活的非特异性内向(主要是是N)离子流,标志符号为1L,的通道允许Na通过,但不同于快Na通 道,两者激活的电压水平不同:1可被艳(Cs)所阻断,而河豚毒却不能阻断它。目前,关于1及其通道的研究资料尚有若干不能充分子以解 释的疑点,对1的进一步研究正受到心肌电生理学者们的高度关注。 然的诗电位及其形机房结含有丰富的 泡,其跨电位有许 作电位复极后出现明显的 ,但它是 种慢反应自律纸 的 浦肯野 )长得 原因 a的 因 动 开文 远不如 后者那 ④没有明显的复极1期和平台期 自动除极速度(约0.1V)却比 野 (约0.02V/s)要快,记录曲线上窦房结细胞4期膜电位变 受化的左别 图4-8心室肌(八)与空房结(B)细狗跨膜电位的此较 窦房结细胞的直径很小,进行电生理研究有一定困难。直到0年代中期,才开始在实房结小标本上采用电压钳技术对其跨膜离子流进行了 定量研究,但目前尚未能充分阐明它的跨膜电位,尤其是4期起搏电流的离子基础。学者们观察到,窦房结细胞0期除极不受细胞外Na浓度的 影响,对河豚毒很不敏感;相反,它受细胞外C✉2+浓度的明显影响,并可被抑制钙通道的药物和离子(如异搏定、D600和M+等)所阻断。 居此可以认为,引起窦房结细胞动作电位0期除极的内向电流是由Ca负载的。这种内向电流被称为第二内向电流:而引起快反应细胞(心室 肌、心房肌和浦情野细泡)》0期除极的快、。*内流称为第一内向电流,根暑已有的研究资料,可将窦房结细抱动作电位的形成过程描述如下:当 摸电位由最大复极电位自动除极达国电位水平时,数话膜上钙通道,引起C:+内向流1ca】,导改0朋除极:随后,钙通道逐渐失话,C+内 流相应减少:另一方面,在复极初明,有一种K*通道被激活,出现K*外向流(4)·C2内流的逐渐减少和K*外流的逐渐增加,膜便逐渐复 极。由“慢”通道所控制、由C2*内流所引起的缓慢0期除极,是变房结细胞动作电位的主要特征,因此,相应称为慢反应细胞和慢反应电位 以区别于前述心室肌等快反应细胞和快反应电位。 窦房结细胞的4期自动除极也由随时间而增长的净内向电流所引起,但其构成成分比较复杂,是几种跨膜离子流的混合,目前已知,在宝房 结细胞4期可以记录到三种提电流,包括一种外电流和两种内向电流,不过它们在窦房结细胞起棒活动中所起作用的大小以及起作用的时间有 所不同
一方面引起另一次动作电位,一方面又反过来中止这种内向电流。这一连串的过程是自律细胞“自我”启动、“自我”发展,又“自我”限制的,由 此可以理解为什么自律细胞能够自动地、不断地产生节律性兴奋。 图4-7 浦肯野细胞起搏机制 A;跨膜电位 B:由х闸门控制的Ik衰减以及由у闸门控制的If,两者在形成起搏电位中的相对关系 这种4期内向电流,通常称为起搏电流,其主要离子成分为Na+ ,但也有K +参与。由于使它充分激活的膜电位为-100mV,因而认为,构成 起搏内向电流的是一种被膜的超极化激活的非特异性内向(主要是是Na+)离子流,标志符号为I f。I f的通道允许Na+通过,但不同于快Na+通 道,两者激活的电压水平不同;I f可被铯(Cs)所阻断,而河豚毒却不能阻断它。目前,关于I f及其通道的研究资料尚有若干不能充分予以解 释的疑点,对I f的进一步研究正受到心肌电生理学者们的高度关注。 2.窦房结细胞的跨膜电位及其形成机制 窦房结含有丰富的自律细胞,动作电位复极后出现明显的4期自动除极,但它是一种慢反应自律细 胞,其跨膜电位具有许多不同于心室肌快反应细胞和浦肯野快反应自律细胞的特征:①窦房结细胞的最大复极电位(-70mV)和阈电位 (-40mV)均高于(电位较正)浦肯野细胞;②0期除极结束时,膜内电位为0mV左右,不出现明显的极化倒转;③其除极幅度(70mV)小于 浦肯野细胞(为120mV),而0期除极时程(7ms左右)却又比后者(1-2ms)长得多。原因是窦房结细胞0期除极速度(约10V/s)明显慢于浦肯 野细胞(200-1000V/s),因此,动作电位升支远不如后者那么陡峭;④没有明显的复极1期和平台期;⑤4期自动除极速度(约0.1V/s)却比浦 肯野细胞(约0.02V/s)要快,记录曲线上窦房结细胞4期膜电位变化的斜率大于浦肯野细胞.图4-8显示心室肌快反应细胞与窦房结细胞跨膜电位 变化的差别。 图4-8 心室肌(A)与窦房结(B)细胞跨膜电位的比较 窦房结细胞的直径很小,进行电生理研究有一定困难。直到70年代中期,才开始在窦房结小标本上采用电压钳技术对其跨膜离子流进行了 定量研究,但目前尚未能充分阐明它的跨膜电位,尤其是4期起搏电流的离子基础。学者们观察到,窦房结细胞0期除极不受细胞外Na+浓度的 影响,对河豚毒很不敏感;相反,它受细胞外Ca 2+浓度的明显影响,并可被抑制钙通道的药物和离子(如异搏定、D-600和Mn 2+等)所阻断。 据此可以认为,引起窦房结细胞动作电位0期除极的内向电流是由Ca 2+负载的。这种内向电流被称为第二内向电流;而引起快反应细胞(心室 肌、心房肌和浦肯野细胞)0期除极的快Na+内流称为第一内向电流。根据已有的研究资料,可将窦房结细胞动作电位的形成过程描述如下:当 膜电位由最大复极电位自动除极达阈电位水平时,激活膜上钙通道,引起Ca 2+内向流(Ica),导致0期除极;随后,钙通道逐渐失活,Ca 2+内 流相应减少;另一方面,在复极初期,有一种K +通道被激活,出现K +外向流(Ik)。Ca 2+内流的逐渐减少和K +外流的逐渐增加,膜便逐渐复 极。由“慢”通道所控制、由Ca 2+内流所引起的缓慢0期除极,是窦房结细胞动作电位的主要特征,因此,相应称为慢反应细胞和慢反应电位, 以区别于前述心室肌等快反应细胞和快反应电位。 窦房结细胞的4期自动除极也由随时间而增长的净内向电流所引起,但其构成成分比较复杂,是几种跨膜离子流的混合。目前已知,在窦房 结细胞4期可以记录到三种膜电流,包括一种外电流和两种内向电流,不过它们在窦房结细胞起搏活动中所起作用的大小以及起作用的时间有 所不同