
第二章细胞机制与认知·23 所不同。 中枢神经系统内的一个少突胶质细胞可围绕几 少突胶质细胞 个轴突形成髓鞘的椭。图2.8表明,少突胶质细雕 可在白质束内缠绕几个临近的轴突形成随稍。如图 所示,外周神经系统内的许旺氏细胞只围绕周围的 ,个神经元轴突形成髓鞘。但二者的作用相似:在 围轴突的髓鞘 一朗飞氏结 轴突周围形成电流绝缘体,从而改变轴突内电流传 递的方式。有髓轴突(还有许多轴突是无髓的)的髓 鞘被结节分隔成若干节段。法国的组织学和解剖学 许旺氏细胞 家Louis-Antoine ranvier最是措述了这一现象,因此 这结节通常被称为朗飞氏结(nodes of Ranvier) 髓鞘的结节部位特化为允许电信号,即动作电何 (action potentials)产生并沿轴突传递的膜结构。因 田绕轴突的髓椭 朗飞氏结 此有髓轴突的髓椭包围部位和无堕鞘的结节部位都 对神经元的信号电流传递具有重要的意义。 当然不 可的部位发挥不同的作用。 图2.8 少突胶质细胞和许旺氏细胞在轴突周围形成 系纺 少突胶质细 的轴突从而形成许多轴突的髓椭,而 周神经系统 的许旺民细胞 烧一根轴突形成静 枢或外周神经 统内,由于每个胶质细 只能理 小段轴突,因此整个轴突髓鞘的形成需要多个胶质细 的参与。 大脑如何工作 髓鞘与疾病 J.C.在32岁时,开始出现走在平坦整洁的车道上 足坐,还可以通过反射系统以保持平衡和姿势,在切 的跑症状。由于对此批的表现感到十分尴尬,她 实生活中 要实现这一过程需要速度和精稀的计的 笑着告诉她的明友那条车道该修整了,然而两周后 因此人从某个角度而言神经系统足一架高速的机器,而 地在穿过客厅的时候跌倒了。在这些笨拙的动作表现 在某些疾病状态下,当神经元及具组成成分的完整性 后的6个月,地又出现了长达6个月之久的一些微 查到破坏时,这一机器的计时精确性将大打折扣 却讨厌的视觉问题,但这些问题在接下来的6个月里 MS是很多类似的神经疾病的一种,这些疾病的 消失了,第一年,】.C又出现了手的麻木和左银的无 共性是特定神经系统内对信交换的协调性出现缺 力症状,为此她不得不去看医生 经过一系列的检查 失 具体而言,MS的病因是中枢神经系统或外周衬 J.C.被确诊为患了一种多发性使化(nultiple sclerosis 经系统(或二者同时)内轴突周围的随稍待异性损伤, MS)的神经肌肉疾病那么这是一种什么样的疾房 通过某种目前尚设有完全了解的机制(可能是某种形 为什么会出现上述的各种症状以及为什么1.C.有时会 式的针对髓鞘本身分子的自身免疫反应),髓鞘被损 出现笨拙的表现? 坏,在少数情况下甚至被完全破坏。髓鞘破坏的程度 在秒水平上,人类的抽经系统时时刻都在加 决定了病人的病情足轻微还是严重。MS病人的临床 工感觉信息以及执行运动反应。其中有些过程是在自 症状则取决于那里的神经元轴突发生脱酷鞘病变。如 主控制下,旧绝大多数的这类加工过程是反射性或自 果病变发生在视觉通路,则出现视觉问题。如果是外 动化的。以站立为例,我们不仅可以自主决定是站还 周神经出现脱精鞘病变,则可能出现感觉、肌肉控制

24·认知神经科学 能力和力量的缺失。如果脱髓鞘病 变发生在涉及脑高级功能的大脑皮 快速信号传递 质连接区的白质束,将可能出现认 划和人格的双变。 为什么精销病变能弓引起这些问 题?髓鞘受损能导致神经信号传递 减慢甚至完全阳断,因此造成受其 完整鞘鞘 影防的地经环路的对应邮分功能缺 失。此外,院髓鞘病变的炎症也能 引起轴突本身的破坏.因此M5的 症状具有两个病因:脱髓销和神经 元受损。随鞘在轴突传导中的作用 将在课文中讨论。在阅读完神经元 (a)健康神经元: 信号传递的内容后,尝试推论髓鞘 (b)多发性硬化病 的同新性损伤如何虑MS的症状。 MS损伤的髓鞘 人的神经元。 神经信号 通过回顾神经元形态学方而的内容,我们注意 理形式的信号(如皮肤躯体感觉感受器接收的触摸 到,神经元的功能是分析和传递信息,这也是神经系 眼睛光感受器接收的光线电突触的电信号等)。这 统的基本日标。接下来要讨论的是神经元之间如何实 些信号引起突出后神经元细胞膜的变化,这种变化导 现信号传递的。 致电流流入或流出神经元。电流在神经元内发挥信号 神经元发放信号需要以下几个必各条件。与自然 作用,并潜在地彬响远离传人突触位点的神经元膜。 世界的大多数事物一样,能量是主要的必备条件。神 电流是由离子流传导的,离子流载有带电荷的离子, 经元如何为神经元发放信号提供能量?能量如何用 如溶于神经元内外液体的钠、钾和氯。这些离子流与 于神经元内信号的产生?神经元间如何实现信号的相 电流相关但不同于金属电线内的电流,后者是由电了 互传递?本节内容将回答这些问题并学习如何通过记 而不是溶解的离子传导的。 录神经元的电活动来研究它们的物理特性。首先是概 长程传递的信号,即动作电位,是在一些被称为 论,然后是具体内容」 锋电位启动区(spike--)的区域产生的 这些区蜮整合来自许多突触传入的或者来自被刺激的 神经元信息交流概论 感受器的电流。锋电位在此特指为动作电位。在大彩 神经元是构成神经系统的完整功能单位,每个神 数情况下,动作电位的结果是一个沿轴突下行传播到 经元通常通过位于树突或胞体的突触来实现与其他 轴突末梢的信号,在那里,最终引起突触神经递质的 神经元的信息交流。传出信息是通过轴突末梢的突触 释放。 实现的。神经元加工信息的目的是接收信息,评估信 本节内容者重阐述的是神经元信号发放的经典 息,然后将信号传递给其他神经元,由此构成了局部 方式,但实际上神经系统内存在着多种形式的传导信 或长程神经环路。 号。例如,许多神经元间的信息交流并不产生动作电 神经元的信号加工包括以下几个阶段。首先神经 位。这些神经元包括位于视网膜内的神经元以及其他 元接收信号,包括化学形式的信号(神经递质,也可 通过电突触相互联系的神经元。 能是环境中能够产生感觉的化学成分,如气味)、物
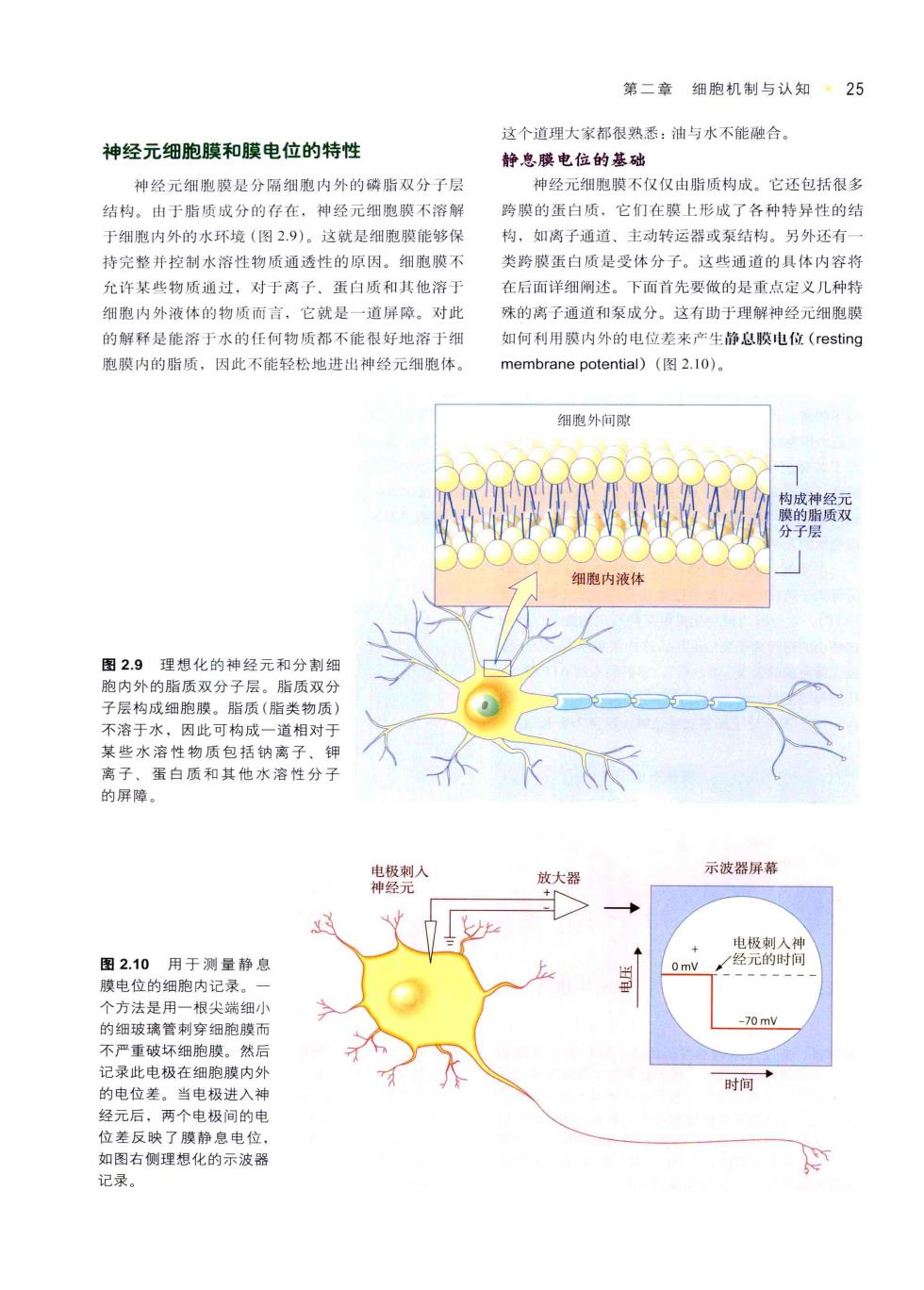
第二章细胞机制与认知·25 这个道理大家都很熟悉:油与水不能驻合 神经元细胞膜和膜电位的特性 静息膜电位的基础 神经元细胞膜是分隔细胞内外的磷脂双分子层 神经元细胞膜不仅仪由脂质构成。它还包括很多 结构。由于脂质成分的存在。神经元细胞膜不溶解 骑膜的蛋白质,它们在膜上形成了各种特异性的结 于细胞内外的水环境(图2.9)。这就是细胞膜能够保 构,如离子通道,主动转运器或泵结构。另外还有 持完整并控制水溶性物质通透性的原因。细胞膜不 类跨膜蛋白质是受体分子。这些通道的具体内容将 允许某些物质通过,对于离子、蛋白质和其他溶于 在后面详细闸述。下面首先要做的是重点定义几种特 细胞内外液体的物质而言,它就是一道屏障。对此 殊的离子通道和泵成分。这有助于理解神经元细胞膜 的解释是能溶于水的任向物质都不能很好地溶于细 如何利用限内外的电位差来产生静息膜电位(resting 胞膜内的脂质,因此不能轻松地进出神经元细胞体 membrane potential)2.10). 细胞外句 神 细胞内液体 图2.9 理想化的神经元和分制 内外的脂质入 质双 因此可构 适相对 性物 质包括到 离 蛋白质和其他水溶性分 屏障 经 示波器屏幕 放大黑 图2.10用于测量静息 模电位的细胞内记灵, 个方法是用一根尖端细小 的细玻璃管刺穿细胞膜而 -70mY 不严重破坏细胞膜。然用 记录此电极在细胞膜内 时间 的电位差。当电极进入刘 经元后,两个电极间的电 位差反映了膜静息电位 如图右侧理想化的示波器 记录

26·认知神经科学 离子通道(ion channel)由跨膜蛋白质所构成, 泵到细胞内同时将3个Na离子泵到细胞外(图2.11), 这些蛋白造成一些孔,这些孔是跨膜通道,能允许 泵的重复话动改变了神经元细胞内一外的K+和Na 钠.钾和氯离子(Na,K和CI)穿过(图2.11) 的浓度,从而产生了膜内外的离子浓度梯度图(图 数以千计的离毛通首存在手神经元细胞膜之中。其中 2.12b)。如果遵循渗透的自然法则,这些离子将从高 行些离子通道是被动的(无佩门的),即水远保持 浓度区域流向低浓度区城,但神经元细胞膜的相对不 个状态(只对某些离子开放)。而另外一些离子通道 可渗透性倾向于阻止这些穿膜的离子移动。因此,在 则是由闸门控制的,即可在电、化学或物理刺激下开 静息状态,神经元细胞膜外是较高浓度的Na,而细 放或关闭。我们将在涉及动作电位产生的内容中具体 胞内则是较高浓度的K·。 阐述后者。 细胞膜的选择性渗透与跨膜的离子浓度梯度的建 离子通道允许离子穿过细胞膜的程度被称为渗透 立导致了细胞膜内外的电位差。具体方式如下:泵建 性(permeability)。尽管细胞膜本身相对面言不允许 立这样的浓度梯度:细胞外Na浓度较高,细胞内K 离子渗透,但一些离子通道(无闸门控制)则允许离 浓度较高。这一梯度将产生一种驱动力,即离子分布 子进出细胞膜。细胞膜对某些离子(如K)的渗透性 不平衡造成的驱动力。这一驱动力试图将Na从高浓 高于其他离子(如N和C),这被称为选择性渗 度区推向低浓度区(即从细胞外到细胞内)以及K从 透性。神经元细胞膜对K'渗透性高于N的原因是 高浓度区推向低浓度区(即从细胞内到细胞外)。但由 膜内的无闸门控制的K通道远多于Na通道(或其 于细胞膜对K的选择性渗透高于Na,这种浓度梯 他类型的无门通道) 主动转运因子如Na-KATP酶泵(图2.I2a)】 a创 细外 能将离子跨膜转运。能量存储分子被称为三磷酸腺省 (ATP),它能够为神经元提供某种形式的能量以维持 这些小的跨膜离子泉的运作。这些泉实际上是位于神 经元细胞膜的酶类(蛋白质),它们能水解ATP分子, 从而释放能量,将Na运出细胞外并将K运进细胞 内。每个ATP分子能够提供足够能量将2个K离子 ADP+(P. 细胞外 细胞内液 [Na]NarK系 [c-] 细胞外 无通的 INa [A-] (c [K+] 细胞内液 细胞内液 图2.11神经元膜内的离子通道。每种离子通道和 图2.12(a)钠-钾泵通过保持细胞内的高浓度K 泵对应不同种类的离子。代表离子的字母周围的方括 和细韵外的高沈度阳而维持细响静白由位复使田 号是浓度的标志,方括号内字母的字体大小与该离子 ATP作为能量。(b)细胞膜对某些离子的选择性渗透 在神经元内外的浓度高低相对应。例如,神经元外的 和主动离子泵所形成的离子浓度梯度导致了跨膜的电 N浓度高于神经元内。神经元细胞膜的特性和穿膜 位差,被称为静息膜电位。该膜电位是神经元细胞外 的主动离子泵共同决定了Na,K和C~及大的载有 正电荷数目多于细胞内,这是图2,10的跨膜电位差的 电荷的蛋白质(Aˉ)的阶梯状浓度 理论基础

第二章细胞机制与认知 27 度的服动力使得一些K+流出细胞外。结果又导致 Bernstein的肌肉组织的测量结果与Nemst方程 另一种照动力 -电荷梯度(electrical gradient)的出 式的计算结果非赏一致。对于神经元,静意草电位主 现,原因在于流出细胞外的K*携带一个正电荷,因 要等于K的平衡电位。而由于其他离子的存在,实 此神经元细外的环境有更高的正电性。 际测量值有所不同。K'并不是细胞膜所通透的唯 对干K+而言,这两种梯度(电荷和离子浓度)被 离子,因此必须考虑Na和C1.20世纪40年代 此是相反的。当带有正电荷的K‘离开细胞时,细胞 David Goldman推导出了一个方程式,该方程式充分 内的不断增强的负电荷环境将阻碍K·离开(正负相 考虑了所有相关离子通透性的计算,被称为Goldman 吸的原理)。最终推动K向细胞外流动的浓度梯度驱 方程式。 动力和将K拉回细胞内的电荷梯度驱动力将达到 20世纪的30年代至50年代,Alan Hodgkin和 个平衡点。这就是两种相反的驱动力所达到的电化学 Andrew Huxley及其学生,同事对Nemst和Goldman 平衡。 方程式开展了经典的模拟检验工作。他们完成了一系 在此平衡状态下的膜两侧微弱的电位差就是静息 列的巧妙研究,包括观察人为地控制细胞内外离子浓 膜电位的基础。如图20所示,跨膜电位可用电极 度对膜电位的影响。他们还在记录膜电位时注入电流, 进行测量。在平衡状液下,这一电位差是-40至-90 观察碳电位的变化并确定相关的穿膜电流。事实上 毫伏(mV) 即神经元细胞内电位负于细胞外电位40 他们的研究结果表明 方程式的确能很好地解释他们 至90毫伏。就此意义而言,神经元与电池一样,它 所记录的膜电位改变。关于他们的新记录方法的内容 且有某种形式的申位能量,通主动的细胞膜过得可 详风“锁如何工作:动作电位历史” 改变细膜电位, 从而使神经元产生电信号 这将在 后而的章节详细讨论。 神经元导电性 膜电位的数学物理模型 对于细胞信号而言,神经元具有两个重要的特 神经元细胞膜的选择性渗透和离子的浓度差导致 性。首先,神经元是容积导体,因此电流可以从其 了静息膜电位。静息膜电位的值是可量化的,19世 内部通过,也可以穿过它们的膜。其次,它们能生 纪德国科学家Walter Nemst提供了方法。他的Ncms 成多种电流包括感受器电位(re eptor potential) 方程式源自物理化学和热力学法则 描述了可溶性 突触电位(synaptic potential)和动作电位(action 子不均衡的分布如何形成渗诱膜(人工膜或生物组织 polential)。在深人讨论神经元的被动电特性之前。 膜)的电位差。该方程式提供了同种离子参与的电位 先来阐述如何测量神经元内的电流和穿膜的电流。 的计算方法.20世纪初Julius Bemstein用此方程式来 膜电位记录 解决生物组织如肌肉如何产生电学势的问题。Nernst 在20世纪40年代和50年代早期,用于高度精 的方程式帮助Bemstein认识到细胞内外离子浓度的 确地记录神经元膜电位的技术手段得以实现。尽管神 差异导致了神经元跨膜电位的差异,K的Nemnst方 经元膜电位测量的基本原理和工具并不复杂,但由于 程式是 钟经元的直径只有50um~100um而轴突直径只有 E=(R2n1og.(K1/[K]D μm 一3μm,因此要实现此记录需要新的技术。为 其中E<代表K的平衡电位,R是普适气体常数,T 了记录膜两侧的电位差异,必须有两个非常小的记录 是凯尔文温标的绝对温度,z是离子的价态(K·是 电极(细线或微电极),一个在细胞内,一个在细 I),F是法拉第常数,[K。是细胞外的K+浓度, 外,两个电极之间的电位差就是跨膜电位差(图2.10) [K]是细胞内的K浓度:1og。是取自然对数的运 为制作一个玻璃微电极,我们需要将一根小玻璃 算。如果将神经细胞内K*浓度400mM和细胞外K 管加热并拉伸成一个细细的尖端(1以m)。当此微电 浓度20mM分别代人方程式,可以将25℃时的K的 极被充入导电溶液时,它就同时扮演了记录电极和束 Nemnst方程式筒化为 穿神经元的刺针的角色。细胞膜内外的电极记录到的 Ex=59.81og,20/400)=-75mV 电压的差异信号,以被放大并在示被器屏幕上显示 这就是K'的平衡电位(equilibrium potential)。需要 出来以供观察】 一般神经元细胞外被定义为零电位 强调指出的是,这是在离子净流量为零时的膜电位。 这样神经元内的静息电位是负电位