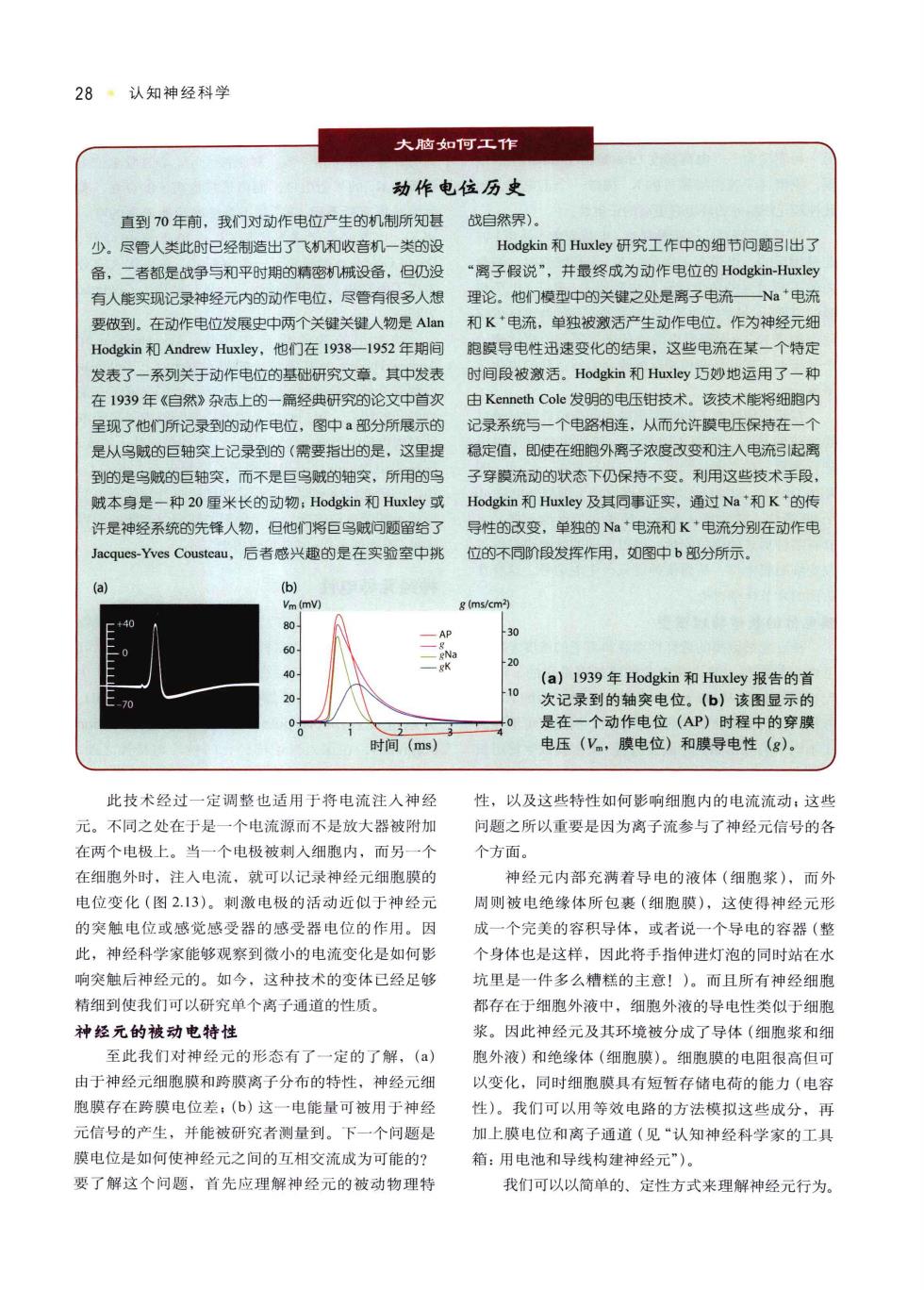
28·认知神经科学 大物和回工作 动作电位历史 直到70年前,我们对动作电位产生的机制所知层 战白然界) 少尽管人类此时已经制造出了飞机和收音机一类的 Hodgkin和Huxley研究工作中的细节问题引出了 备,二者都是战争与和平时期的精密机械设备,但仍没 “离子假说”,并最终成为动作电位的Hodgkin-Huxley 有人修实现记录神经元内的动作电位,尽管有很多人想 理论。他们模型中的关键之处是离子电流一 一Na电开 要放到。在动作电位发展史中两个关键关键人物是A 和K+电流,单独被激活产生动作电位。作为神经元细 Hodgkin和Andrew Huxley,他门在1938一1952年期间 胞膜导电性迅速变化的结果,这些电流在某一个特发 发表了一系列关于动作电位的基础研究文章。其中发表 时间段被激活。Hodgkin和Huxley巧妙地运用了一种 在1939年《自然》杂志上的一篇经典研究的论文中首 由Kenneth Cole发明的电压钳技术。该技术能将细跑内 呈现了他门所记录到的动作电位,图中a部分所展示的 记录系统与一个电路相连,从而允许模电压保持在一 是从鸟诚的巨轴突上记录到的(需要指出的足,这里提 稳定值,即使在细胞外离子浓度改变和注人电流起离 到的是鸟威的巨轴突,而不是巨鸟贼的轴突,所用的 子穿膜流动的状态下仍保持不变。利用这些技术手段 赋本身是一种20里米长的动物:Hodgkin和Huxley或 Hodgkin和Huxley及其同事证实,通过Na和K的代 许是神经系统的先锋人物,日他门巨岛城问颗留给了 导性的改变,单独的Na电流和K电流分别在动作用 s-Yves Cousteau 后者感兴趣的是在实验室中桃 位的不同阶段发挥作用,如图中b部分所示 my) e (ms/cm? -A (a)1939年Hodgkin和Huxley报告的首 20 次记录到的轴突电位。(b】该图显示的 是在 个动作电位(AP)时程中的穿膜 时间(ms) 电压(V,膜电位)和膜导电性(g)。 此技术经付一定调整巾话用干将电流注人神经 性,以及这些特性如何影响细胞内的电流流动:这些 元。不同之处在于是一个电流源而不是放大器被附加 问题之所以重要是因为离子流参与了神经元信号的各 在两个电极上。当一个电极被刺入细胞内,而另一个 个方面 在细胞外时,注入电流,就可以记录神经元细胞膜的 神经元内部充满着导电的液体(细胞浆),而外 电位变化(图2.13)。刺激电极的活动近似于神经元 周则被电绝缘体所包裹(细胞膜),这使得神经元形 的突触电位或感觉感受器的感受器电位的作用。因 成一个完美的容积导体,或者说一个导电的容器(整 此,神经科学家能够观察到微小的电流变化是如何影 个身体也是这样,因此将手指伸进灯泡的同时站在水 响突触后神经元的。如今,这种技术的变体已经足够 坑里是一件多么糕的主意!)。而且所有神经细 精细到使我们可以研究单个离子通道的性质 都存在于细胞外液中,细跑外液的导电性类似于细胞 神经元的被动电持性 浆。因此神经元及其环境被分成了导体(细胞浆和细 至此我们对神经元的形态有了一定的了解,(a) 胞外液)和绝缘体(细胞膜)。细胞膜的电阻很高但口 由于神经元细胞膜和跨膜离子分布的特性,神经元细 以变化,同时细胞膜具有短暂存储电荷的能力(电容 胞膜存在跨膜电位差:(b)这一电能量可被用于神经 性)。我们以用等效电路的方法模拟这些成分,耳 元信号的产生,并能被研究者测量到 个问题是 加上膜电位和离子通道(见“认知神经科学家的工具 膜电位是如何使神经元之间的互相交流成为可能的? 箱:用电池和导线构建神经元”)。 要了解这个问题,首先应理解神经元的被动物理特 我们可以以简单的、定性方式来理解神经元行为
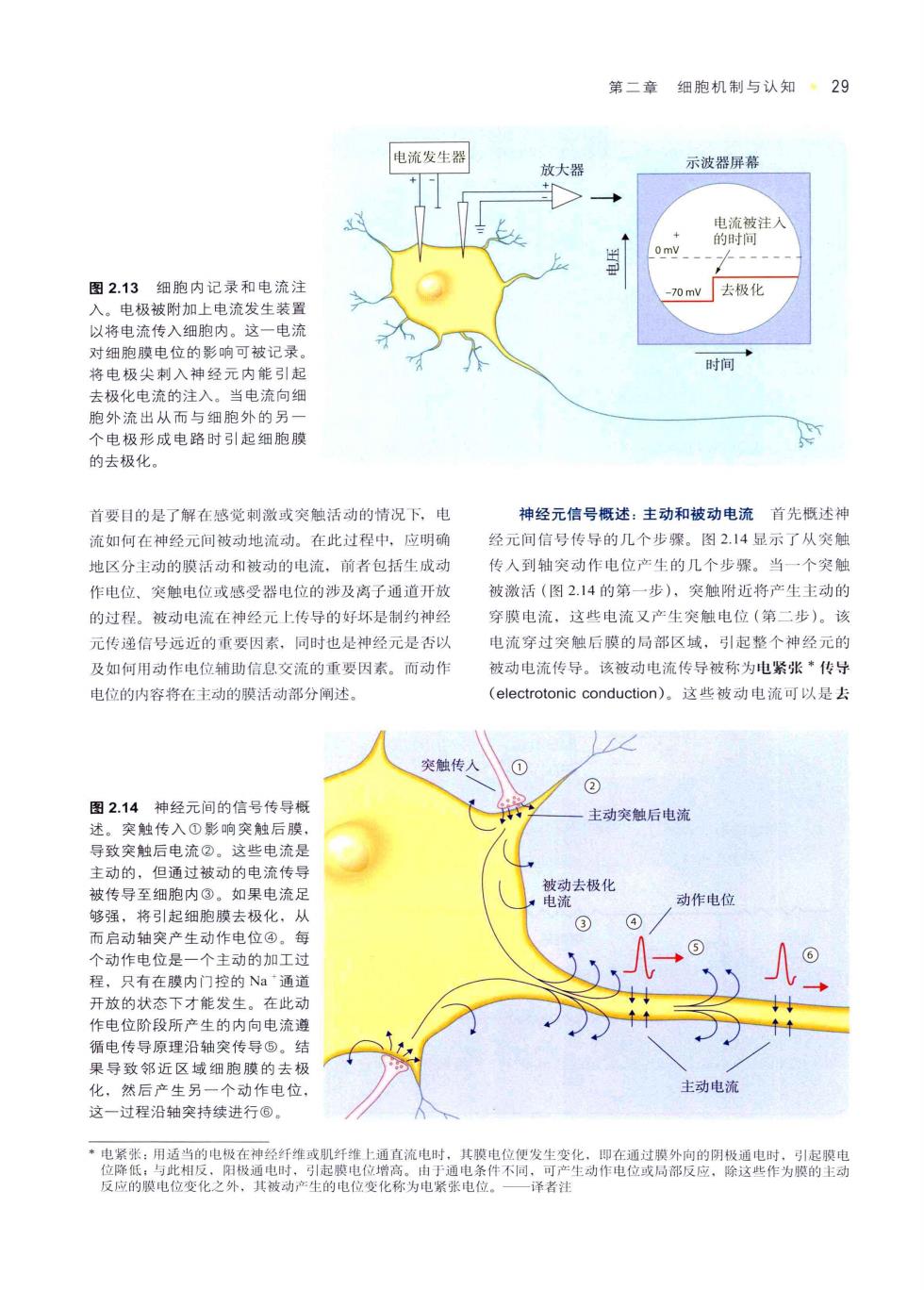
第二章细胞机制与认知·29 电流发生器 放大器 示波器屏幕 图2.13细胞内记录和电流注 0mv去极化 入。电极被附加上电流发生装置 以将电流传入细胞内。这一电流 时细胞膜申位的影响可被记最 将电极尖刺入神经元内能引起 时间· 去极化电流的注入。当电流向细 跑外流出从而与细胞外的另 个电极形成电路时引起细胞膜 的去极化。 首要目的是了解在感觉刺激或突触活动的情况下,电 神经元信号概述:主动和被动电流首先概述相 流加何在神经元间被动地流动。在此过程中,应明确 经元间信号传导的几个步零。图2.14显示了从突制 地区分主动的膜活动和被动的电流,前者包括生成动 传入到轴突动作电位产生的几个步骤。当一个突 作电位、突触电位或感受器电位的涉及离子通道开放 被激活(图2.14的第一步),突触附近将产生主动的 的过程。被动电流在神经元上传导的好坏是制约神经 穿装电流,这些电流又产生突触电位(第二步)。该 元传递信号远近的重要因素。同时也是神经元是否以 电流穿过突触后膜的局部区域,引起整个神经元的 及如何用动作电位辅助信息交流的重要因素。而动作 被动电流传导。该被动电流传导被称为电紧张*传号 电位的内容将在主动的膜活动部分阐述 (electrotonic conduction)。这些被动电流可以是去 ∠ 突触传人 图2.14 神经元间的信号传导 主动突触后电流 突触 但通过被 电流传 电流 ,去极化 动作电位 够强 动作电位是 只有在膜内 控的N 的内向查纯 理滑 过程沿轴突持续进行© 电紧张用当的申在神经仟维或肌 通直流电时 译者注
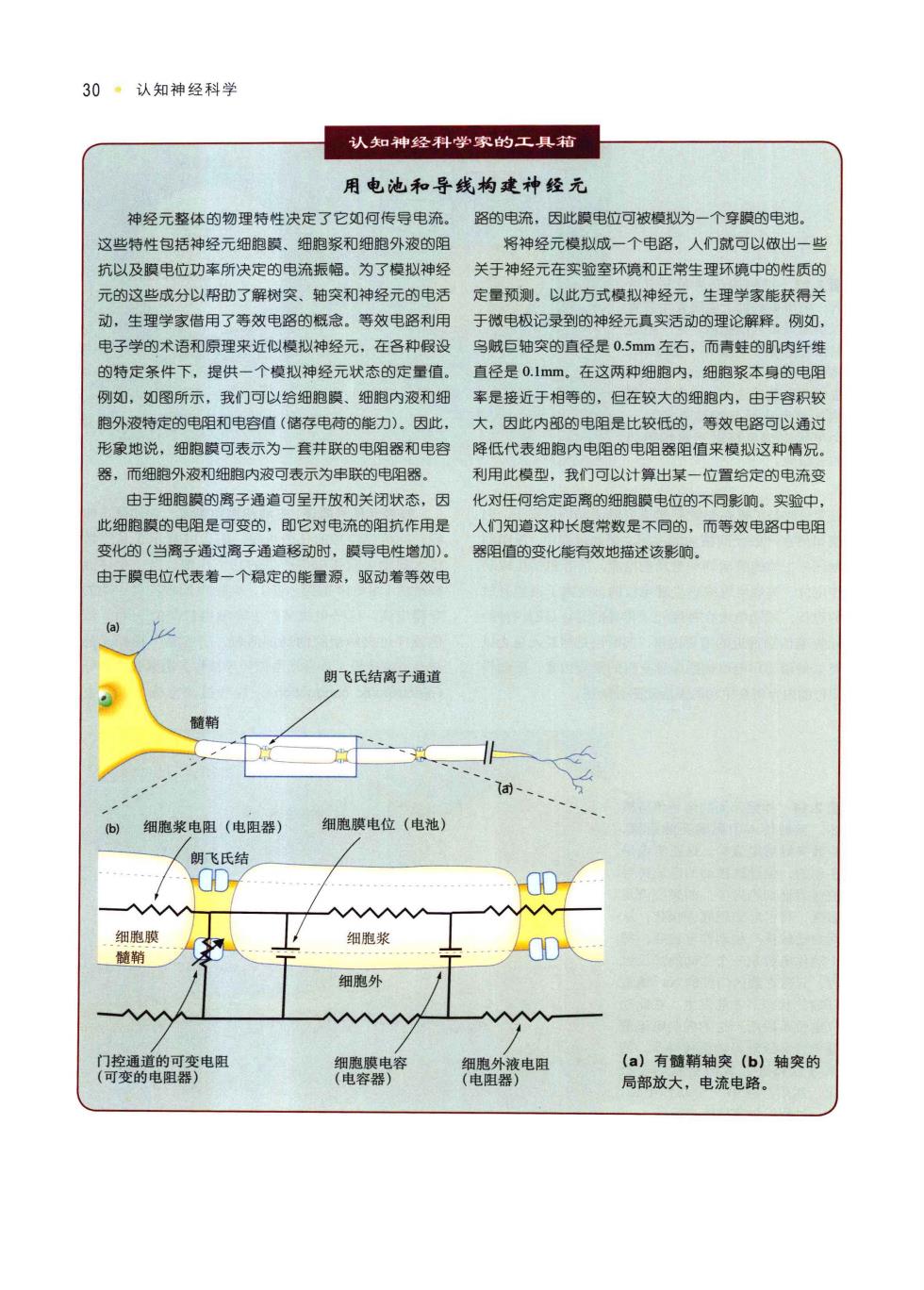
30。认知神经科学 认知神经科学家的工具箱 用电池和导线构建神经元 神经元整体的物理特性决定了它如何传导电流, 路的电流,因此膜电位可被模拟为一个穿膜的电池, 议些特性包括神经元细胞膜、细胞浆和细胞外液的阳 将神经元模以成一个电路,人们就可以做出一些 抗以及膜电位功率所决定的电流振幅。为了模拟神经 关于神经元在实验室环镜和正常生理环境中的性质日 元的这些成分以帮助了解树突、轴突和神经元的电活 定量预测,以此方式模拟神经元,生理学家能获得关 动,生理学家借用了等效电路的概念,等效电路利用 于微电极记录至到的神经元真实活动的理论醒降、网如」 电子学的术语和原理来近似模拟神经元,在各种假衫 鸟贼巨轴突的直径是0,5mm左右,而青蛙的肌肉纤维 的特定条件下,提供一个模似神经元状态的定量信。 直径是0.1mm。在这两种细胞内,细胞浆本身的电阻 列测如,如图所示,我门可心以给细报膜细胞内液和细 案是接近于相等的,旧在较大的细胸内,中干容积敏 胞外液特定的电阻和电容值(储存电荷的能力)。因此 大,因此内部的电阻是比较低的,等效电路可以通过 形象地说,细胞膜可表示为一套并联的电阻器和电容 降低代表细胞内电阻的电阳器阻值来模拟这种情况 器,而细跑外液和细胞内瘦可表示为虫联的电阻器 利用此模型,我门可以计出其一竹置给定的电森 由于细胞装的离子通道可呈开放和关闭状态,因 化对任何给定更离的细胞膜电位的不同影响。实验中 此细胞膜的电阻是可变的,即它对电流的阻抗作用是 人门知道这种长度常数是不同的,而等效电路中电阳 变化的(当离子酒过离子通消超动时.膜号电性增加) 器阳值的变化能有效地描述该影前 由于摸电位代表着 一个稳定的能量源,驱动着等效甲 朗飞氏结离子通道 细胞浆电阻(电阻器 细胞膜电位(电池) D) 朗飞氏结 AAAA 细胞浆 细胞外 AAAAA 电容 电 (a)有髓精轴突(b)轴突的 局部放大,电流电路

第二章细胞机制与认知·31 极化(depolarization)的(被称为兴奋性突触后电位) 是小离子,因此可被称为穿膜的Na流,K流和C 使细胞内的电环境更趋正性,从而更容易产生动作 流,或若一般地说是细胞内或外的离子流。在化学 电位:也可能是超极化(hyperpolarization)的(被利 电或物理刺激下,细胞膜离子通道开放,Na和K 为抑制性突触后电位)。使细胞内的电环境更趋负性 穿膜流动,从而形成电流 从而不容易产生动作电位 电紧张(衰减)传导理解神经元信号传导的 被动电流通过细跑浆的容积导体被传导通过突 个关健问题是:在没有主动过程(即动作电位)补充 触后神经元的树突和胞体(第三步)。如果这些被过 能量的情况下,被动离子电流沿树突、轴突或胞体传 电流能有效地使神经元去极化,它们将在此神经元 导的距离有多远?该距离取决于神经元的三个主要物 轴丘的锋电位启动区激发动作电位(第四步)。动作 理特性:()起始电流的幅度,(b)神经元细胞膜的电 电位是主动的膜活动(原因在于它涉及的膜导电性改 阻(和电容):()细胞内液和细胞外液的导电性。通 变是通过离子通道开放和关闭实现的),但就结果而 过对神经元的模拟研究和直接的实验观察,如记录神 言,就像在突触附近一样,它所导致的结果是轴突 经元内电活动(对于足够大的细胞)及将电流注入神 内产生被动电流(第五步)。这些被动电流又引起邻 经元以观察结果(图2.16),我们已逐渐开始理解这 近的轴案区域去极化,引起新的动作电位产生(第六 些基本原理。这些方法较详细地阐述了被动的膜特性 步),此过程沿轴突不断进行,直至轴突末精。然 以及它们如何参与神经元信号的活动 此末梢释放神经递质,上述过程将在下 个神经元 首先,考虑图2.17a所示的神经元 一个刺入祺 全部“上演” 经元的刺澈电极可被用于将电流注入细内来榄拟突 有时很难准确地了解动作电位所引起的主动电 触传入的活动 然后在轴突上距离该刺激电极刺入 如何能导致被动电流。当流入神经元的电流处于发生 的不同距离处插入记录电极。当制激电极将正性电清 主动膜活动的附近区域时,它被称为主动电流(后面 注入该轴突部位时(即电极尖端是正性的),电流 的章节将详细述)。同样的电流接着通过电紧张传 轴突传递,在轴突膜上表现出来 并形成返回电流 导而被动地流过细胞时,我们称它们为被动电流。这 流向位于细胞外液中的另一个电极。这样形成一个完 是因为当它们流过细真时,它们不是通过离子通道的 整的电路。在此图例中,注入电流所诱发的去极化程 开放而传导 而是通过细胞膜的物理特性和电路的 度低于产生动作电位的阀值(见下一节)。刺激电极 性传导的。一个完整的电路要求电流必须特续地流 附近的电流强度是最强的。被动电流以递减方式沿射 动,因此离子的细胞内流动必定伴随着电流向细雕外 突传递。如果记录穿膜电压值,侧大的电压变化出现 返回,从而形成一个完整的电路(图2,15)。 在刺激电极附近,而距离越远,电压值变化越小。如 如前所述,神经元电流是离子流,也就是说它们 果假设细胞膜的电阻是不变的,则膜电压值化的 是由溶液中带有电荷的原子(离子)承载的。神经元 原因在于电流幅度随距离增加而衰减(即信号递减) 的电荷载体主要是Na'、K和C,也包括一些溶 因此电紧张传导也被称为衰诚传导。 液中的带有电荷的大分子蛋白质。由于穿膜电流通常 轴突内和轴突周闲电流传递方式类似于声音、手 电流发生器 5动作电位产生的 穿膜电流,沿神经元 个与电流发生 出神经元 图216人工模拟诱导的电流。利用 重回到起始部 器相连的细胞内电极,可将电流注入神经元

32·认知神经科学 (al 电流发生器 100 电紧张传导 看起始值的 电位 记录电极 距电流注入点的距离 电脉冲 图2.17 (a)电极在神经元轴突的 一端注入电流,记录电极位于不同距离的轴突位点。 由注入电流所诱发的 低于产生动作电位阔值的膜的去极化振幅随距离 6) 沿轴 的下睡 传的始部位至膜电 与距离延长呈指数关系,已知的长度常数()值被定义为从膜电位变 值37%处的距离。 电筒光柱或匹萨饼气味的传递,都是随着距离增加而 变化是分级的 逐渐减弱。欧姆定律(电压=电流×电阻,或V= 那么细胞膜电阻对电位传导性有何影响?简单 1×R)可帮助我们理解这种关系。当电流沿轴突流动 地说,当题电阻提高时,电流将以在轴突内流动为 时,累积的电阻(R)越来越大,电路中的电压(V)随 主,而很少流出。设想一下情况与此相类似的公园 之下降的幅度也越来越大。我们可以将此方程式改写 管道系统。如果管道系统的水管是完好的(即对水的 为1=VR,表示当电阻增加时,电流强度()下降。 穿透力具有高阻抗),则位于水管一端的水将被迫在 然而应该指出的是,欧姆定律的这种简单表达式,不 水管内流动。而如果管道内遍布孔隙(对水的阻抗 能完全说明神经元的各种细节特性,尽管它对建立 低),则水管一端的水在流动过程中不断地外漏,水 关于电紧张传导的相关因素的感性认识很有帮助(见 流的流动距离变短。如果管道系统的渗漏到了一定 “认知神经科学家的工具箱:用电池和导线构建神经 程度,水流的流动距离将变得非常短。同样,神经 元")。 元细胞膜的电阻能影响电流在轴突的传导距离(相关 神经元内电传导的目的是传导信号变化,这种信 内容将在髓鞘与传导性的部分进行讨论) 号变化如果足够强,可以引起轴突末梢释放神经递 最后,电阻的倒数一 一细胞内的电导 也能影 质,实现神经元间突触传递。目前最让人感兴趣的问 响神经元电流的传导距离。作为盐溶液,细胞内液和 题是被动电流传导是否足够实现这种传递。答案是, 细胞外液通常是电流的良导体,具有很高的电导(星 因情况而定。由于电流强度随距离增加而衰减,因此 然它们比金属导线还差很多)。即使这样,树突、轴 被动电流传导只在短距离发挥作用,而不适用于长距 突和胞体的电阻随大小不同而变化。再回到公园曾 离信号交流。 道系统的类比,如果水管的直径较大,则水流动相 如前所述,电流传导的有效距离部分依颗于电流 容易,但如果水管较窄,水流受限,水压和阻抗将增 的强度。电流越强,则传导距离越远。在实验条件 大。同样,如果轴突直径较大,则电流流动较远。因 下,可以通过提高注入电流的强度来人为地操纵电流 此, 高幅度的感受器或突触电流,高的细胞膜电阻和 传导的程度。在正常生理条件下,电流幅度由以下生 低的细胞内电阻将增强电流的电传导性,能影响距离 理因素决定,如感受器感受到某一物理刺激的强度以 传人点远处的神经元细胞装」 及神经元上突触传入的强度和数目。由于这些事件可 在最佳条件下,电紧张传导作为电交流方式,其 能与不同强度的电流相关,因此它们所诱发的藻电位 有效性能维持到多远的距离?距离不长,一般只有