
18认知神经科学 第一查向读老措绘了一幅现代认知神经科学的历 生物事件决定行为的准确方式的理解仅仅处于起始 史画卷,闲述了心理学和生物学经过怎样的融合才诞 阶段,但目前的研究已经积系了大量的神经元水平 生了这门新的交叉学科。有关心理学和神经科学历 的生物学知识。其中一些发我可以帮助我们很好地 的理论,思拟和特点在这一章得以体现。尽管有关意 理解脑科学的基本原则。本章将主要回顾这些神经 识与脑的理论可以追溯到古代文明,但目前认为关于 科学的某本知识,为以后章节的学习莫定基。本 神经系统的基本生物学知识主要开始于19世纪下半 章的主要内容包括神经元的细胞解剖学和生理学 叶。神经科学在过去的100年中的发展为理解人的脑 识,第三章将要阐述的内容是神经系统的整体解留 细胞和神经环路如何决定行为奠定了基础。 和发展的知识。为了更好地理解信息加工的生物学 当代神经科学家已理解了神经信号 突触传递 知识 本章将提出以下几个问题:神经元之间如何实 以及信息存储和加工的神经编码等基本概念。研究 现信总交流?作为信息交流媒介的化学信号包括哪 人员致力于在基本的分千及细胞水平的加工与单个 此?药物如何通讨影血神经元闲的交互作用从而对 神经元及神经环路的活动之间建立联系。尽管关于 机体发挥有益或有害的作用 德经系统的细胞 科学家们常采用一种还原主义的策略,希望能通 神经系统由两种细组成。经元和经胶细 过首先确定局部的性质进而了解整体。例如,在过 胞。除了与其他细胞的共同点之外,神经元还具有独 去400年的物理和化学理论中,对构成物质的基本元 特的形态和生理学特性以完成其特有的功能。胶质细 素的探寻一直热火朝天地进行者。这种探寻导致了两 胞是神经系统内的一类非神经元细胞,它承担着多种 方面的重大发现:构成所有分子的基本元素和亚原子 功能,包括对神经元提供结构支持和绝缘以保证神经 结构原理。早期的生物学家也对这种构成我们周遭世 元间信总传递更为有效等。 界的基本部件而着迷。在17世纪和18世纪,他们尝 试用放大镜捕捉生物组织结构的形象。功夫不负有心 神经元结构 人,细跑理论因此而证生细跑理论假完,牛物体是 神经元(neuron)是基本的信号处理单位。它们 由基本单位或细胞所构成的。而正如第一章所指出 依靠各自的外形、功能、位置和神经系统内的相互连 的,单个细胞构成了神经系统的基本功能单位这一理 接方式来彼此区分(图2.I)。正如Ramon y Cajal及 论在20世纪初受到了挑战。最终,Ramon y Cajal的 其间时代的科学家所推断的,神经元接收信息,遵份 神经系统由单个神经元构成的理论被普遍接受, 某些相当简单的规律做出“决定”,然后改变自己的 Ramon y Cajal观察到,尽管神经元彼此之间紧 活动水平,最后将信息传递给其他神经元。这些功能 密相邻,但被很小的缝儆分隔。通过这些观察,他确 与神经元的形态或结构的特异性密切相关 定了神经元的两个主要原理: 神经元的组成成分之一是细胞体(图22。与其 1,连接的特异性:神经元的细胞浆彼此并不相通 他细胞一样,细胞体内包含有维特神经元新陈代谢的 因此每个神经元是相互独立的,而神经元问的这 细胞器。这些细胞器包括细胞核、内质网。核糖体 种连接并不是随意的、它是神经元传递信息的特 线粒体、高尔基器以及其他与大多数细胞相的细胞 异性通路。 器。这些结构被神经元的双层脂质细胞膜所包围,并 2.功能性两授分化,神经元的一部分专门负贵接收 悬浮在细胞质中,细胞浆是存在于所有细胞内流动的 信息,而另一部分则专门负责将信息传递给其他 液体 神经元或肌肉」 除细胞体外,神经元的另一个组成成分是延伸 这两个基本原理和神经元学说的整体理论将是本 至胞体外的特异性突起,树突(dendrites)和轴实 章要刚述的第一部分内容,它将描述神经系统的细胞 (axo)。这两类突起的作用反映了功能两极分化的 结构和功能。 基本原理。树突通常是大树样的突起,它接收来自
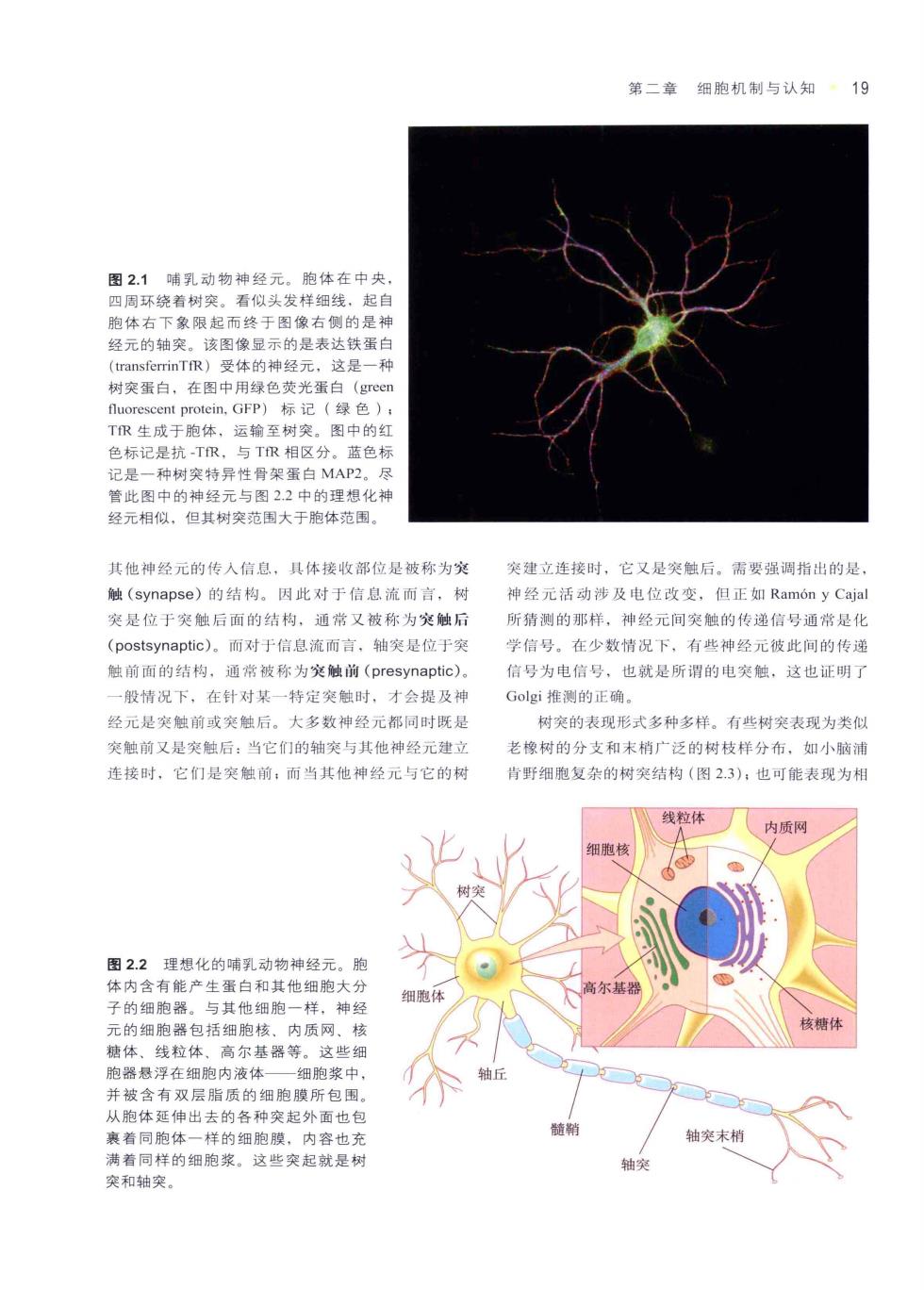
第二章细胞机制与认知19 看似头发样细线。起自 胞体右 起而终于图像右侧的是神 经元的轴突 该图像显示的是表达铁蛋白 树突蛋白,在图中用绿色荧光蛋白(grc TR生成于胞体,运输至树突。图中的红 色标记是抗TR,与TR相区分。蓝色标 记是一种树突特异性骨架蛋白MAP2。尽 管此图中的神经元与图2.2中的理想化神 经元相似,但其树突范围大于胞体范围, 其他神经元的传入信息。具体接收部位是被称为突 突建立连接时,它又是突后。需要强调指出的是 触(synapse)的结构。因此对于信息流而言,树 神经元活动涉及电位改变,但正如Ram6 n y Caia 突是位于突触后面的结构,通常又被称为突触后 新猜测的那样,神经元间突触的传递信号通常是化 (postsynaptic),而对于信息流而言,轴突是位于突 学信号。在少数情况下,有些神经元彼此间的传递 触前面的结构,通常被称为突触前(presynaptic) 信号为电信号,也就是所谓的电突触,这也证明了 般情况下,在针对某一特定突触时, 才会提及神 Gogi推测的正确 经元是突触前或突触后。大多数神经元都同时既是 树突的表现形式多种多样。有些树突表现为类似 笑触前又是突触后:当它们的轴突与其他神经元建立 老橡树的分支和末梢“泛的树枝样分布,如小市浦 连接时,它们是突触前:而当其他神经元与它的树 背野细胞复杂的树突结构(图2.3),也可能表现为相 线粒体 内质网 图2.2理想化的哺乳动物神经元。胞 体内含有能产生蛋白和其他细胞大分 子的细胞器。与其他细胸一样,神经 元的细胞器包括细胞核、内质网、核 糖体、线粒体、高尔基器等。这些细 胞器悬浮在细胞内液体 细胞浆中 并被含有双层脂质的细胞膜所包围, 从胞体延伸出去的各种突起外面也包 裹着同胞体一样的细胞膜,内容也充 轴突末梢 满着同样的细胞浆。这些突起就是树 突和轴突
20·认知神经科学 图2.3小脑浦肯野细胞的胞体和树 树突 轴究 突。浦肯野细胞排列在小脑内。每个 细胞的树突呈现为逐级分支 级太 奔贺曹野相 于一级,最终形成一个庞大的树突 “树” 当简单,如脊髓运动神经元或丘脑神经元的树突(图 轴突末梢具有特异化的细胞内结构,它们使得信息 2.4)。树突亦可具有被称为棘(spine)的特异化突起, 交流通过神经递质的释放而实现,神经递质是神经 即树突表面的细颈连有小的球状突起:突触通常位于 元间化学突触传递信号的化学物质。神经元的树突 这些棘上(图2.5)。突触还存在于神经元的其他部位 和轴突是飞体的延伸都分内部都充满着细浆。因 包括没有棘的细胞体 此,胞体、树突和轴突是属于同一个神经元的不同 细胞体的另一类突起是远离胞体的轴突。该结 组成部分。神经元各组成部分间的细胞内空间的连 构代表着神经元的输出端:电信号传送至轴突末梢 续性对于神经元的电信号功能是必要的,这点将在 即突触所在的位置。正如前面所指出的,这些轴突 以后的内容中得以证明。 和神经元的轴突末梢对于突触而言属于突触前结构。 图2.2所描绘的理想化神经元是以脊髓运动神经 元为模型,而实际上神经元具有多种形式。神经元形 态的多样性令人印象深刻,关于人类神经系统的众名 说明远超过本章篇幅。早期的解剖学家详细描述了这 种形态的多样性并根据它们的形状将神经元分为3利 或4种大的类型。这些分类方法主要根据树突和轴突 相对于彼此或胞体的方向形态。图2.6显示了带见的 4种类型神经元:单极、双极、假单极和多极神经元 形态学相似的神经元倾向于集中在神经系统的某一特 有区域,并具有相似的功能 但神经系统各区城的神 经元形状知千差万别。 一般而言,单极神经元只有一个远离胞体的突 起:此突起能分支形成树突和轴突末梢,此类神经元 常见于无脊椎动物的神经系统。双极神经元主要参 与感觉信息加工,例如,双极神经元传递听觉、视 觉和嗅觉系统的信息。此类神经元具有两个突起: 轴突(至肌肉) 根树突和一根轴突。从某种意义上讲,它可以被看 做原型神经元:通过树突接收来自某一端的信息,通 图2.4前角运动神经元。位于脊髓的多级神经元 过轴突将信息传至另一端,例如眼祝网膜的双极衬 其轴突穿出脊髓前根直至与肌肉纤维建立突触联系 经元(图1,13)。眼视网腴的双极神经元只局限在视
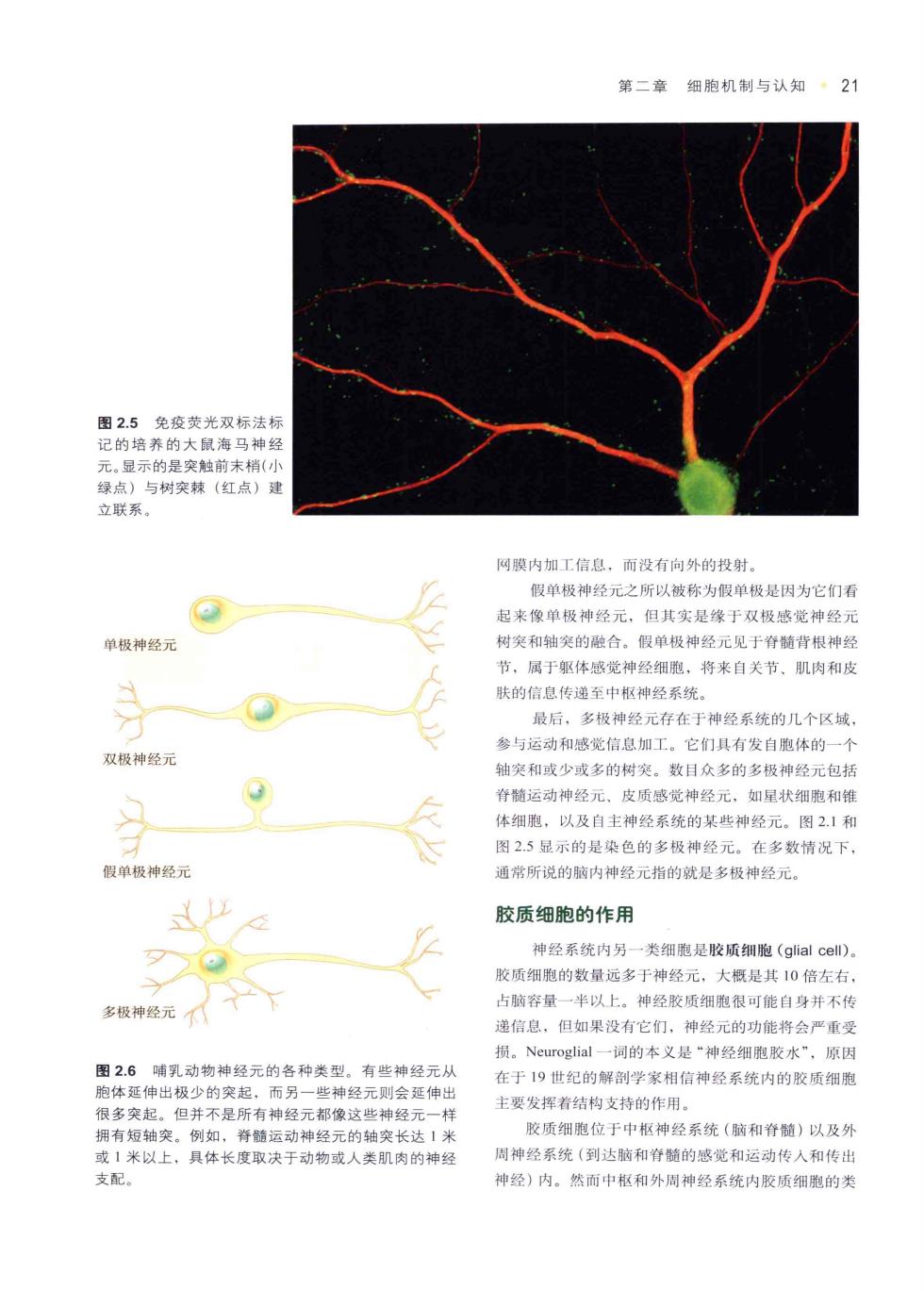
第二章细胞机制与认知·21 图2.5免疫荧光双标法标 神 树突树 立联系 网膜内加工信息,而没有向外的投射 假单极神经元之所以被称为假单极是因为它们看 起来像单极神经元,但其实是缘于双极感觉神经元 单极神经元 树突和轴笑的融合。假单极神经元见于脊髓背根神经 节,属于躯体感觉神经细胞,将来自关节,肌肉和皮 y 肤的信息传递至中枢神经系统 最后,多极神经元存在于神经系统的几个区域 参与运动和感觉信息加工。它们具有发自胞体的一个 双极神经元 轴突和或少或多的树笑。数目众多的多极神经元包括 脊髓运动神经元、皮质感觉神经元,如星状细胞和倒 体细胞,以及自主神经系统的某些神经元。图21和 图25显示的是染色的多极神经元。在多数情况下, 假单极神经元 通常所说的脑内神经元指的就是多极神经元。 以 胶质细胞的作用 神经系统内另一类细胞是胶质细胞(glial cell)。 胶质细胞的数量远多于神经元,大概是其10倍左右, 多极神经元产 占脑容量一半以上。神经胶质细胞很可能自身并不传 递信息,但如果没有它们,神经元的功能将会严重受 损。Neuroglial一词的本义是“神经细胞胶水”,原因 图2.6哺乳动物神经元的各种类型。有些神经元从 在于19世纪的解剖学家相信神经系统内的胶质细胞 胞体延伸出极少的突起,而另一些神经元则会延伸出 王要发挥着结构支持的作用。 很多突起。但并不是所有神经元都像这些神经元一样 拥有短轴突。例如,脊髓运动神 胶质细胞位于中枢神经系统(脑和脊精)以及外 经元的尖长达不 1米以上,具体长度取决于动物或人类肌肉的神经 周神经系统(到达脑和脊髓的感觉和运动传入和传出 支配。 神经)内。然而中和外周神经系统内胶质细胞的类

22·认知神经科学 型是不同的。中枢神经系统内主要有3种类型的胶 伤的组织显微分析表明小胶质细胞大量侵入该受损部 质细胞,早形胶质细胞。少突胶质细胞和小胶质细胞 位。此时小胶质细胞发挥巨游细胞作用,吞噬和清除 (图2.7)) 受损的脑细胞。小胶质细胞在成年人中(与其他胶质 早形胶质细胞是一种呈圆形或放射对称形状的大 细胞一样)能够增殖,而传统认为中抠神经系统的利 细南,它们围终着神经元并与脑血管紧密连接。星形 经元是不可增生的(但最新的研究表明,成年脑也可 胶质细胞与脑血管的接触部位特化为终足 该结构呀 出现神经元再生,参见第三章) 允许离子进出血管壁又在中根报神经系统的组织与血液 神经系统的胶质细胞最主要的功能是构成神经 之间物辣了一道屏隐。这种神经组织与血被之间的异 元的髓鞘。髓销(mvelin)是包绕许多神经元轴突的 形胶质细胞屏障被称为血脑屏障(blood-brain barier 脂类物质。在中枢神经系统内,少突胶质细胞构成酷 BBB)。该屏障能阳挡某些血液传播的病原或过度影 箱:在外周神经系统内,许旺氏细胞担当此功能(图 白神经活性的化学物质的排人,从而在保护中拟神经 2.7)。这两种类型的胶质细胞在神经元的生长发有过 系统中发挥至关重要的作用。例如,许多药物不能等 程中,将它们的细胞膜以同心方式缠绕轴突,并将出 过BBB,某些神经激活因子如多巴胺和去甲肾上腺 处胶质细胞的细胞浆挤出,从而形成髓鞘。细胞浆被 素一旦位于血液中,也不能通过BBB。 挤出,主要留下了磷脂双分子层,这使得随鞘具有特 BBB的这种 不可通透性对于某些疾病的治疗具 征性的脂类物质外观(在正常生理情况下,由于白色 有重要的意义,如帕金森氏症,一种由于基底节多巴 脂类的存在,髓鞘看起来是发亮的白色)。但如表21 胺缺失所导致的严重运动障碍疾病(见第三章和第七 所示,少突胶质细胞和许旺氏细胞形成髓鞘的方式有 章),病因在于产生多巴胺并通过其轴突将多巴胺运 送至基底节的黑质神经元的变性丢失。由于多巴胺不 表2.1形成髓鞘的两种神经胶质细胞的 能通过BBB,因此直接通过血液注射多巴胺并不能 主要差异 补充脑内衰竭的多巴胺。但血液中的多巴胺生物合成 的前体物质分子L-Dopa却能穿过BBB,从而被神经 细胞类型 位置 元摄取以转化为脑组织的多巴胺。 许旺氏细胞 外周神经系统 小胶质细胞是一种形状小而不规则的神经胶质 少突胶质细胞 中枢神经系统 多个 细胞,主要在脑组织损伤时发挥作用(图27)。脑损 早形细 中枢神经系统 少突胶质细 小胶质细 图27哺乳动物中枢和外周神经系统 内的胶质细胞类型。星形细胞与血管 外周神经系统 的接触部位形成终足。少突胶质细胞 和许旺氏细胞分别构成中枢神经系统 和外周神经系统内神经元轴突的髓鞘。 亦可见小胶质细胞