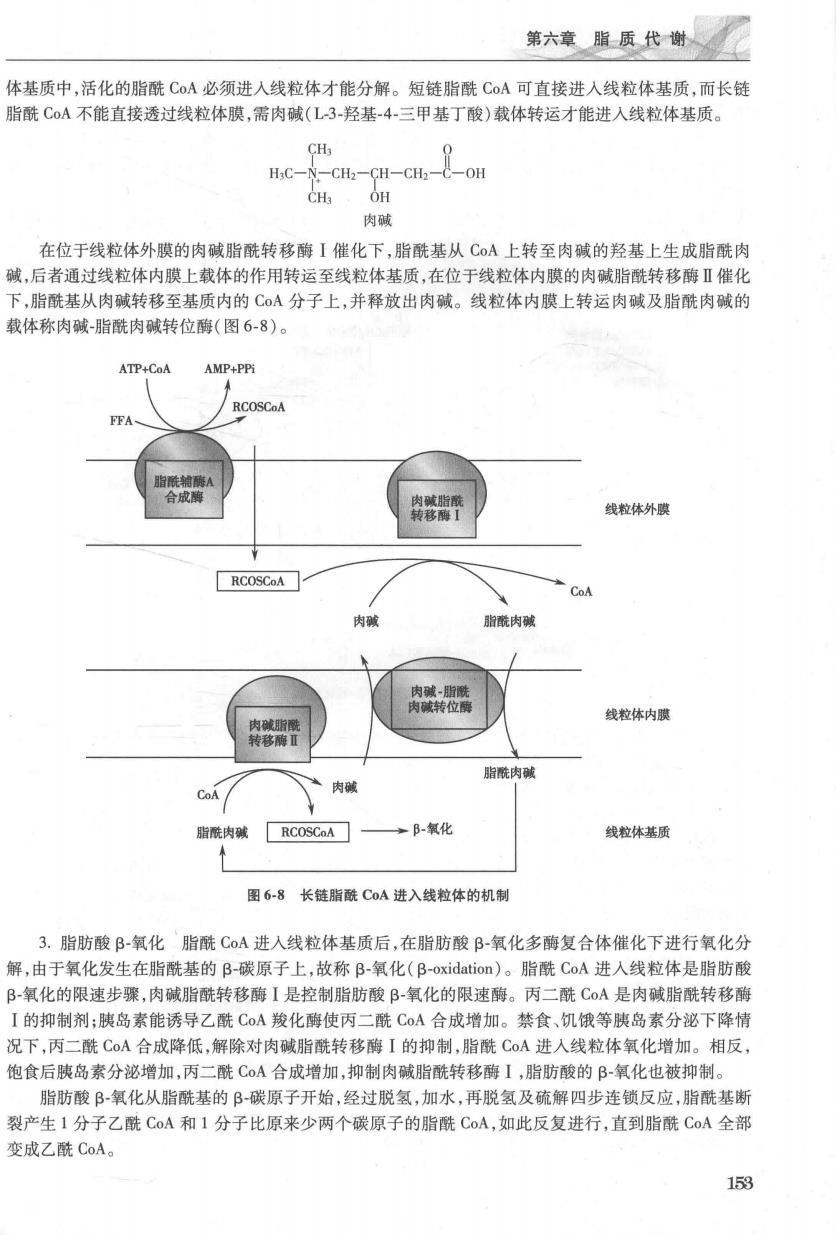
第六章脂质代谢 体基质中,活化的脂酰CoA必须进入线粒体才能分解。短链脂酰CoA可直接进入线粒体基质,而长链 脂酰C0A不能直接透过线粒体膜,需肉碱(L-3-羟基-4-三甲基丁酸)载体转运才能进入线粒体基质。 H:C-N-CHz-CH-CH2-C-OH CHs OH 肉碱 在位于线粒体外膜的肉碱脂酰转移酶【催化下,脂酰基从C0A上转至肉碱的羟基上生成脂酰肉 碱,后者通过线粒体内膜上载体的作用转运至线粒体基质,在位于线粒体内膜的肉碱脂酰转移酶Ⅱ催化 下,脂酰基从肉碱转移至基质内的CA分子上,并释放出肉碱。线粒体内膜上转运肉碱及脂酰肉碱的 载体称肉碱-脂酰肉碱转位酶(图6-8)。 ATP+CoA AMP+PPi 脂酰酶A 合成酶 线粒体外膜 RCOSCoA CoA 肉或 脂酰肉碱 肉 肉碱脂酰 线粒体内膜 转移酶Ⅱ 指脂酰肉碱 CoA 肉碱 脂酰肉碱厂RCOSCoA门 →B-氧化 线粒体基质 图6-8长链脂酰C0A进入线粒体的机制 3.脂肪酸β-氧化脂酰CoA进入线粒体基质后,在脂肪酸B-氧化多酶复合体催化下进行氧化分 解,由于氧化发生在脂酰基的B碳原子上,故称B-氧化(B-oxidation)。脂酰CoA进人线粒体是脂肪酸 β氧化的限速步骤,肉碱脂酰转移酶I是控制脂肪酸B-氧化的限速酶。丙二酰C0A是肉碱脂酰转移酶 I的抑制剂;胰岛素能诱导乙酰C0A羧化酶使丙二酰CoA合成增加。禁食、饥饿等胰岛素分泌下降情 况下,丙二酰C0A合成降低,解除对肉碱脂酰转移酶I的抑制,脂酰C0A进入线粒体氧化增加。相反 饱食后胰岛素分泌增加,丙二酰C0A合成增加,抑制肉碱脂酰转移酶I,脂肪酸的B-氧化也被抑制。 脂肪酸B氧化从脂酰基的β-碳原子开始,经过脱氢,加水,再脱氢及硫解四步连锁反应,脂酰基断 裂产生1分子乙酰CoA和1分子比原来少两个碳原子的脂酰CoA,如此反复进行,直到脂酰CoA全部 变成乙酰CoA。 153
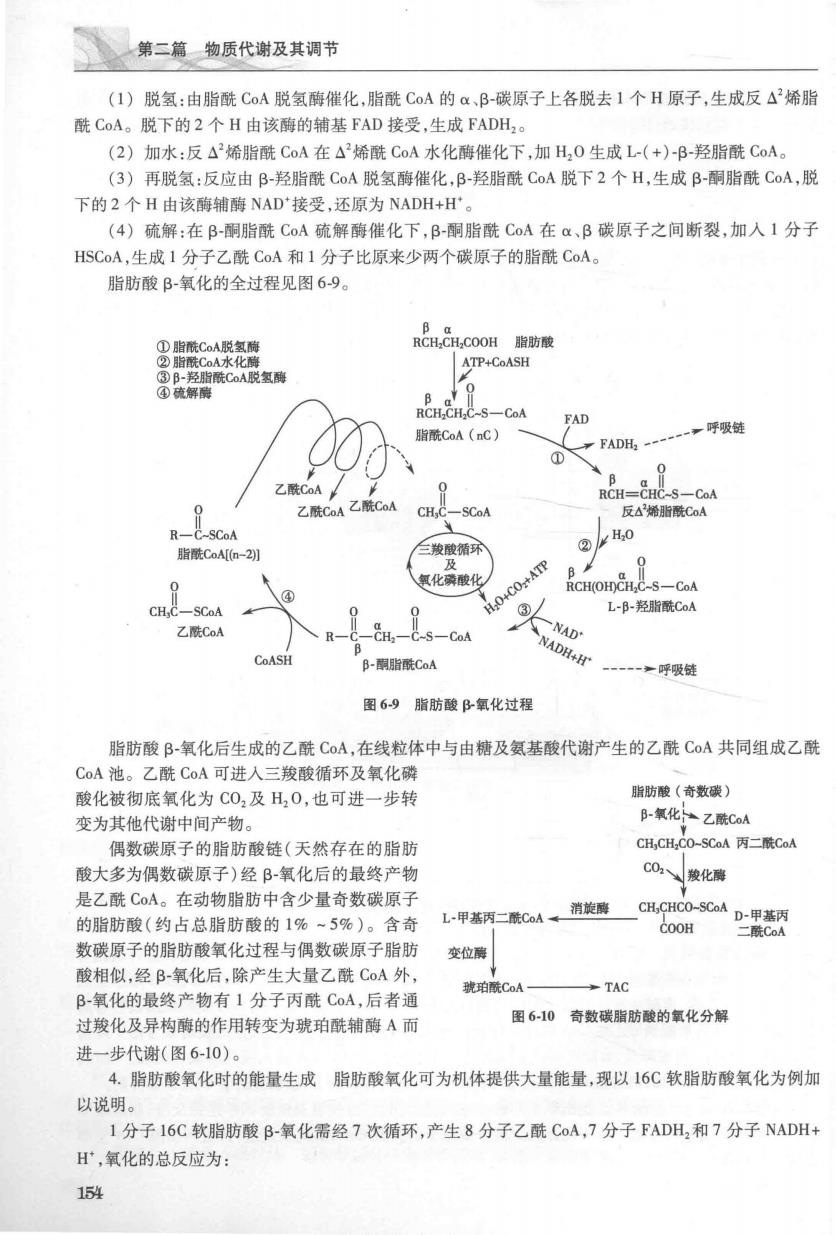
第二有类质代游及其调节 (1)脱氢:由脂酰CoA脱氢酶催化,脂酰CoA的α、B-碳原子上各脱去1个H原子,生成反△烯脂 酰CoA。脱下的2个H由该酶的辅基FAD接受,生成FADH2。 (2)加水:反△2烯脂酰CoA在△烯酰CoA水化酶催化下,加H,0生成L-(+)-B-羟脂酰CoA。 (3)再脱氢:反应由B-羟脂酰CoA脱氢酶催化,B-羟脂酰CoA脱下2个H,生成B-酮脂酰CoA,脱 下的2个H由该酶辅酶NAD接受,还原为NADH+H'。 (4)硫解:在B-酮脂酰C0A硫解酶催化下,B-酮脂酰CoA在α&、B碳原子之间断裂,加入1分子 HSCA,生成1分子乙酰CoA和1分子比原来少两个碳原子的脂酰CoA 脂肪酸B-氧化的全过程见图69。 ①脂酰C。A脱氢稀 RCH,CH.COOH脂肪酸 ②脂酰CoA水化酶 ATP+CoASH 多幕反氧 ROCA FAD 脂酰CoA(nC) 一,呼吸链 FADH2- 0 乙酰CoA 乙酰CoA乙酰CoA RCH= IC-S-CoA CHC-SCoA 反△烯胎酰C0A R-C-SCoA 脂酰CoAn-2别 三羧酸循环 及 氧化磷酸化 RCH(OH)CHC-S-CoA ④ CH;C-SCoA H.C ③4 L-B-羟脂酰CoA 乙酰COA CH2-C-S-CoA NAD CoASH B-酮脂酰CoA NADH+H 一呼吸链 图69脂肪酸B氧化过程 脂肪酸B-氧化后生成的乙酰C0A,在线粒体中与由糖及氨基酸代谢产生的乙酰CoA共同组成乙酰 CoA池。乙酰C0A可进入三羧酸循环及氧化磷 酸化被彻底氧化为C0,及H,0,也可进一步转 脂肪酸(奇数碳) 变为其他代谢中间产物。 B-氧化乙酰CoA 偶数碳原子的脂肪酸链(天然存在的脂肪 CH,CH,C0-SCoA丙二酰CoA 酸大多为偶数碳原子)经B-氧化后的最终产物 C02骏化碳 是乙酰C0A。在动物脂肪中含少量奇数碳原子 的脂肪酸(约占总脂肪酸的1%~5%)。含奇 L甲背mL男 COOH 数碳原子的脂肪酸氧化过程与偶数碳原子脂肪 变位 酸相似,经B-氧化后,除产生大量乙酰C0A外 琥珀酰CoA →TAC B-氧化的最终产物有1分子丙酰C0A,后者通 过羧化及异构酶的作用转变为琥珀酰辅酶A而 图6-10奇数碳脂肪酸的氧化分解 进一步代谢(图6-10)。 4.脂肪酸氧化时的能量生成脂肪酸氧化可为机体提供大量能量,现以16C软脂肪酸氧化为例加 以说明。 1分子16C软脂肪酸B-氧化需经7次循环,产生8分子乙酰CoA,7分子FADH2和7分子NADH+ H,氧化的总反应为: 54
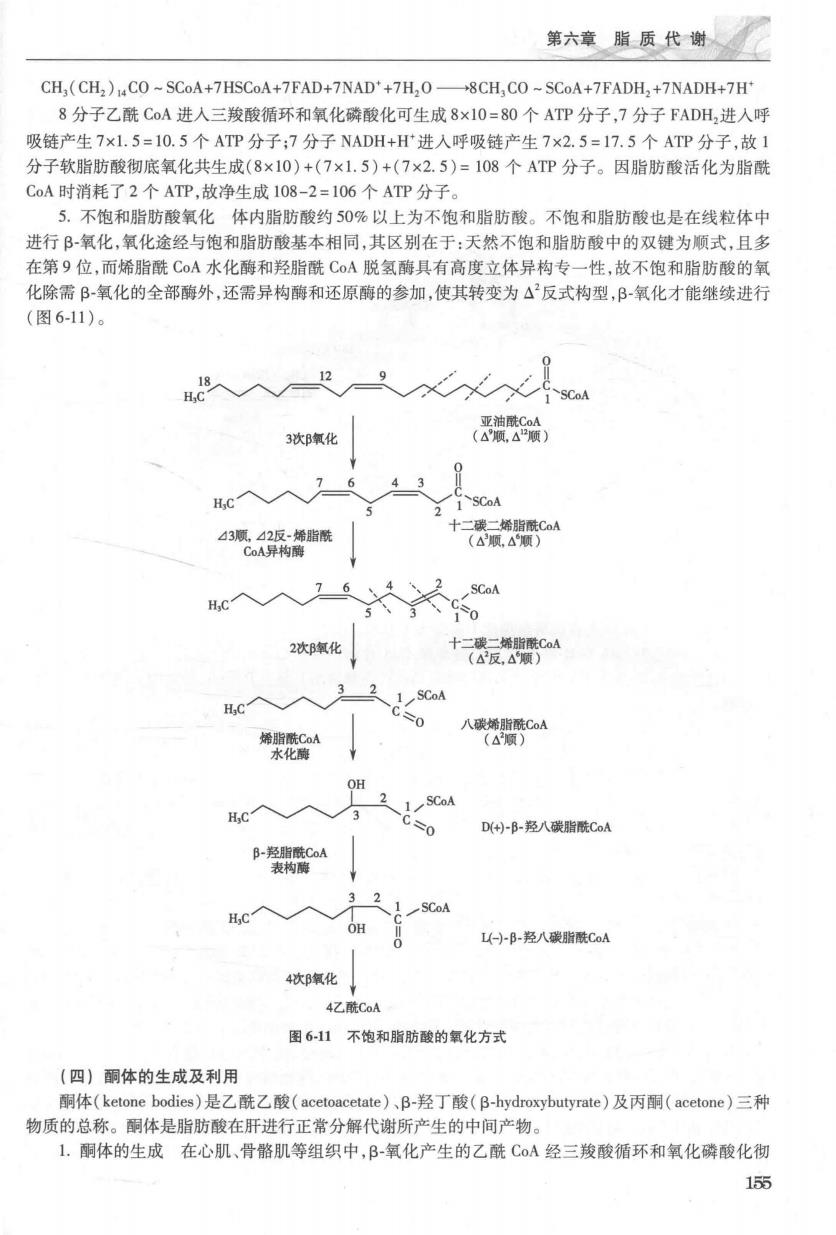
第六章脂质代谢 CH,(CH2 )CO~SCoA+7HSCoA+7FAD+7NAD*+7H,O-8CH,CO-SCoA+7FADH,+7NADH+7H* 8分子乙酰CoA进人三羧酸循环和氧化磷酸化可生成8×10=80个ATP分子,7分子FADH2进人呼 吸链产生7×1.5=10.5个ATP分子;7分子NADH+H进入呼吸链产生7×2.5=17.5个ATP分子,故1 分子软脂肪酸彻底氧化共生成(8×10)+(7×1.5)+(7×2.5)=108个ATP分子。因脂肪酸活化为脂酰 CoA时消耗了2个ATP,故净生成108-2=106个ATP分子。 5.不饱和脂肪酸氧化体内脂肪酸约50%以上为不饱和脂肪酸。不饱和脂肪酸也是在线粒体中 进行B-氧化,氧化途经与饱和脂肪酸基本相同,其区别在于:天然不饱和脂肪酸中的双键为顺式,且多 在第9位,而烯脂酰C0A水化酶和羟脂酰CA脱氢酶具有高度立体异构专一性,故不饱和脂肪酸的氧 化除需B-氧化的全部酶外,还需异构酶和还原酶的参加,使其转变为△反式构型,B氧化才能继续进行 (图6-11)。 3次B氧化 流 7 43 H,C SCoA 432脂酰 十最 异构 H,C入 76 SCoA 0 2次B氧化 十 32 H.C 1 SCoA 90 八碳烯脂酰CoA 烯脂酰CoA 水化 OH 2 C 3 00 D(+)-B-羟八碳脂酰CoA B-羟脂酰CoA 表构酶 SCoA L)B-羟八碳脂酰CA 4次B氧化 4Z酰CaA 图6-11不饱和脂肪酸的氧化方式 (四)酮体的生成及利用 酮体(ketone bodies)是乙酰乙酸(acetoacetate)、B-羟丁酸(B-hydroxybutyrate)及丙酮(acetone)三种 物质的总称。酮体是脂肪酸在肝进行正常分解代谢所产生的中间产物。 1.酮体的生成在心肌、骨骼肌等组织中,B-氧化产生的乙酰CoA经三羧酸循环和氧化磷酸化彻 155