
第十章DNA的生物合成本科六版第十章DNA的生物合成一、基因(gene)1868年瑞士外科医生Miescher从细胞中分离到核酸,1909Johannsen首先提出用基因代表遗传物质。而关于基因的本质直至五十年代后才得以逐渐揭示。定义:DNA分子上编码生物活性产物的功能片段。这些生物活性产物包括蛋白质和RNA。二、中心法则(the centraldogma of molecular biology)DNA上的基因只是一种信息,这种信息只有表达出来才有意义。在生物体内基因表达的具体方式主要是蛋白质。1、1958年Crick基于诸多室验依据提出生物体内遗传信息表达的基本规律一一中心法则:转录翻译复制RNA蛋白质DNA逆转录1970年H.Temin发现了逆转录现象,使中心法则得以扩充。2、复制(replication):以DNA为模板合成DNA的过程。其本质是将遗传信息由亲代准确地传给子代的过程。3、转录(transcription):以DNA如模板合成RNA的过程,即将遗信息由DNA传给RNA的过程。4、翻译(translation):以RNA的模板合成蛋白质的过程,即将遗传信息由RNA传给蛋白质的过程。5、基因表达(geneexpression):指遗传信息由DNA经RNA最终表达为蛋白质的过程。包括转录、翻译两个步骤。6、逆转录(Reversetranscription)以RNA为模板合成DNA的过程。即将遗信息由RNA逆向传给DNA的过程。其遗传信息传递方向与转录相反。故名逆转录。-1
第十章 DNA 的生物合成 本科六版 - 1 - 第十章 DNA 的生物合成 一、基因(gene) 1868 年瑞士外科医生 Miescher 从脓细胞中分离到核酸,1909 Johannsen 首先提出用基因代表遗传物质。而关于基因的本质直至五十年代后才得以逐渐 揭示。 定义:DNA 分子上编码生物活性产物的功能片段。 这些生物活性产物包括蛋白质和 RNA。 二、中心法则(the central dogma of molecular biology) DNA 上的 基因只是一种信息,这种信息只有表达出来才有意义。在生物 体内基因表达的具体方式主要是蛋白质。 1、1958 年 Crick 基于诸多室验依据提出生物体内遗传信息表达的基本规 律——中心法则: 转录 翻译 复制 DNA RNA 蛋白质 逆转录 1970 年 H.Temin 发现了逆转录现象,使中心法则得以扩充。 2、复制(replication):以 DNA 为模板合成 DNA 的过程。其本质是将遗 传信息由亲代准确地传给子代的过程。 3、转录(transcription):以 DNA 如模板合成 RNA 的过程,即将遗信 息由 DNA 传给 RNA 的过程。 4、翻译(translation):以 RNA 的模板合成蛋白质的过程,即将遗传信 息由 RNA 传给蛋白质的过程。 5、基因表达(gene expression):指遗传信息由 DNA 经 RNA 最终表达为 蛋白质的过程。包括转录、翻译两个步骤。 6、逆转录(Reverse transcription)以 RNA 为模板合成 DNA 的过程。 即将遗信息由 RNA 逆向传给 DNA 的过程。其遗传信息传递方向与转录相反。故 名逆转录

第十章DNA的生物合成本科六版本篇以中心法则为主线索,着重阐述遗传信息传递的基本规律(复制、转录、翻译)。在此基础上,介绍基因表达的调控和分子生物学的新技术。DNA的生物合成包括复制和逆转录,复制为大多数生物所共有,而逆转录只存在于个别生物体中,故本章着重讲述与复制相关的内容。第一节复制的方式一一半保留复制当细胞分裂时,DNA即开始复制,其主要特征为半保留复制(semi-conservative replication)。一、半保留复制的定义:DNA的两条链均可作为模板,合成出子代DNA与母代完全相同,且一条链来自母链,一条链是新合成的,这种复制方式称为Semi-conservative replicatior二、半保留复制的实验依据:1953年Watson和Cmick在提出双螺旋模型时即已推测出复制方式。1958年M.Messe1son和F.W.Stahl用实验加以证实。实验方法见P221图11-1三、半保留复制的意义:由于DNA分子中两股链有碱基互补关系,一条链即可确定其互补链的碱基序列。按半保留复制的方式,子代保留了亲代的全部遗传信息,通过基因表达,决定了子代生物的特性和类型。故半保留复制的意义为:保持了代与代之间DNA序列的一致性,体现了遗传过程的相对保守性。第二节参与复制的酶参与复制大分子物质有:1、底物(substrate):dNTP(N:A、T、C、G)2、模板(template):母代DNA的两条单链。3、引物(primer):为一有3一OH的寡核苷酸。4、酶和蛋白:包括聚合酶、解链解旋酶、引物酶、连接酶等。一、DNA 聚合酶- 2 -
第十章 DNA 的生物合成 本科六版 - 2 - 本篇以中心法则为主线索,着重阐述遗传信息传递的基本规律(复制、 转录、翻译)。在此基础上,介绍基因表达的调控和分子生物学的新技术。 DNA 的生物合成包括复制和逆转录,复制为大多数生物所共有,而逆转录 只存在于个别生物体中,故本章着重讲述与复制相关的内容。 第一节 复制的方式——半保留复制 当细胞分裂时, DNA 即开始复制,其主要特征为半保留复制 (semi-conservative replication)。 一、半保留复制的定义:DNA 的两条链均可作为模板,合成出子代 DNA 与母代完全相同,且一条链来自母链,一条链是新合成的,这种复制方式称为 Semi-conservative replication 二、半保留复制的实验依据:1953 年 Watson 和 Cmick 在提出双螺 旋模型时即已推测出复制方式。1958 年 M.Messelson 和 F.W.Stahl 用实验加以 证实。 实验方法见 P221 图 11-1 三、半保留复制的意义:由于 DNA 分子中两股链有碱基互补关系,一 条链即可确定其互补链的碱基序列。按半保留复制的方式,子代保留了亲代的 全部遗传信息,通过基因表达,决定了子代生物的特性和类型。故半保留复制 的意义为:保持了代与代之间 DNA 序列的一致性,体现了遗传过程的相对保守 性。 第二节 参与复制的酶 参与复制大分子物质有: 1、底物(substrate):dNTP(N:A、T、C、G) 2、模板(template):母代 DNA 的两条单链。 3、引物(primer):为一有 3’—OH 的寡核苷酸。 4、酶和蛋白:包括聚合酶、解链解旋酶、引物酶、连接酶等。 一、DNA 聚合酶

第十章DNA的生物合成本科六版全称为DNAdependentDNApolymerese(DDDP),1958年A.Kornberg在大肠杆菌中发现了polI,并在试管内证明了其聚合酶活性(一)DDDP催化的反应较清楚的是原核生物的DDDP。1、5'一3'聚合活性以母链为模板,按碱基互补规律,DDDP在引物3一OH上逐个加上dNTP使新链延长,见P223反应式为(dNMP)n+dNTP→(dNMP)n+1+PPiDDDP的聚合方向只能是5,一3。2、核酸外切酶(exonucleasae)活性:表现为3'-5’外切酶和5'-3"外切酶活性。3”-5”外切酶在复制的校读及损伤修复中起作用,5’-3”外切酶则在引物切除及损伤修复中起作用。(二)种类1、原核生物的DNA聚合酶:E.coli中有PolI、PolII和polIⅢl,三者在细胞中的分子数之比为400:40:20,但活性polIII是PolI的10倍以上,故认为polIII是复制中起主要作用的酶。po1III的分子量约140KD,由10种亚基组成不对称的二聚体(图11-5A),有聚合酶和3'-5外切酶活性,每分钟催化约10'个核苷酸聚合。其中α、&、0组成核心酶,α亚基有5-3’聚合酶活性,8亚基有3’-5’外切酶活性及碱基选择功能,e亚基可能起维系二聚体的作用。两边的β亚基起结合模板的作用。其余的亚基统称为-复合物,具体功能尚在研究中。Po1I的分子量约为109KD,为单一肽链,以a螺旋为主,有A-R共18个α螺旋区(图11-5B),有聚合酶、3”-5’外切酶和5,-3”外切酶活性,每秒钟催化约10个核苷酸聚合。由于Po1I只能催化合成约20个核苷酸,即从模板上脱离,故主要在校读和修复中起作用。在特异蛋白酶作用下,Po1I可被水解为67KD和36KD大小两个片段(图11-5B的F、G螺旋间):大片段又称K1enow片段,具有聚合酶和3,-5,外切酶活性,小片段具有5,-3,外切酶活性。这两个片段均为分子生物学研究的工具酶。- 3 -
第十章 DNA 的生物合成 本科六版 - 3 - 全称为 DNA dependent DNA polymerese(DDDP),1958 年 A.Kornberg 在 大肠杆菌中发现了 polⅠ,并在试管内证明了其聚合酶活性. (一)DDDP 催化的反应 较清楚的是原核生物的 DDDP。 1、5’—3’聚合活性 以母链为模板,按碱基互补规律,DDDP 在引物 3’—OH 上逐个加上 dNTP 使新链延长,见 P223 反应式为(dNMP)n+dNTP (dNMP)n+1+PPi DDDP 的聚合方向只能是 5’—3’。 2、核酸外切酶(exonucleasae)活性:表现为 3’-5’外切酶和 5’-3’ 外切酶活性。3’-5’外切酶在复制的校读及损伤修复中起作用,5’-3’外切 酶则在引物切除及损伤修复中起作用。 (二)种类 1、原核生物的 DNA 聚合酶: E.coli 中有 PolⅠ、PolⅡ和 polⅢ,三者在细胞中的分子数之比为 400: 40:20,但活性 polⅢ是 PolⅠ的 10 倍以上,故认为 polⅢ是复制中起主要作 用的酶。 polⅢ的分子量约 140KD,由 10 种亚基组成不对称的二聚体(图 11-5A), 有聚合酶和 3’-5’外切酶活性,每分钟催化约 105个核苷酸聚合。其中α、ε、 θ组成核心酶,α亚基有 5’-3’ 聚合酶活性,ε亚基有 3’-5’外切酶活性及 碱基选择功能,θ亚基可能起维系二聚体的作用。两边的β亚基起结合模板的 作用。其余的亚基统称为γ-复合物,具体功能尚在研究中。 PolⅠ的分子量约为 109KD,为单一肽链,以α螺旋为主,有 A-R 共 18 个 α螺旋区(图 11-5B),有聚合酶、3’-5’外切酶和 5’-3’外切酶活性,每秒 钟催化约 10 个核苷酸聚合。由于 PolⅠ只能催化合成约 20 个核苷酸,即从模 板上脱离,故主要在校读和修复中起作用。 在特异蛋白酶作用下,PolⅠ可被水解为 67KD 和 36KD 大小两个片段(图 11-5B 的 F、G 螺旋间):大片段又称 Klenow 片段,具有聚合酶和 3’-5’外切 酶活性,小片段具有 5’-3’外切酶活性。这两个片段均为分子生物学研究的 工具酶
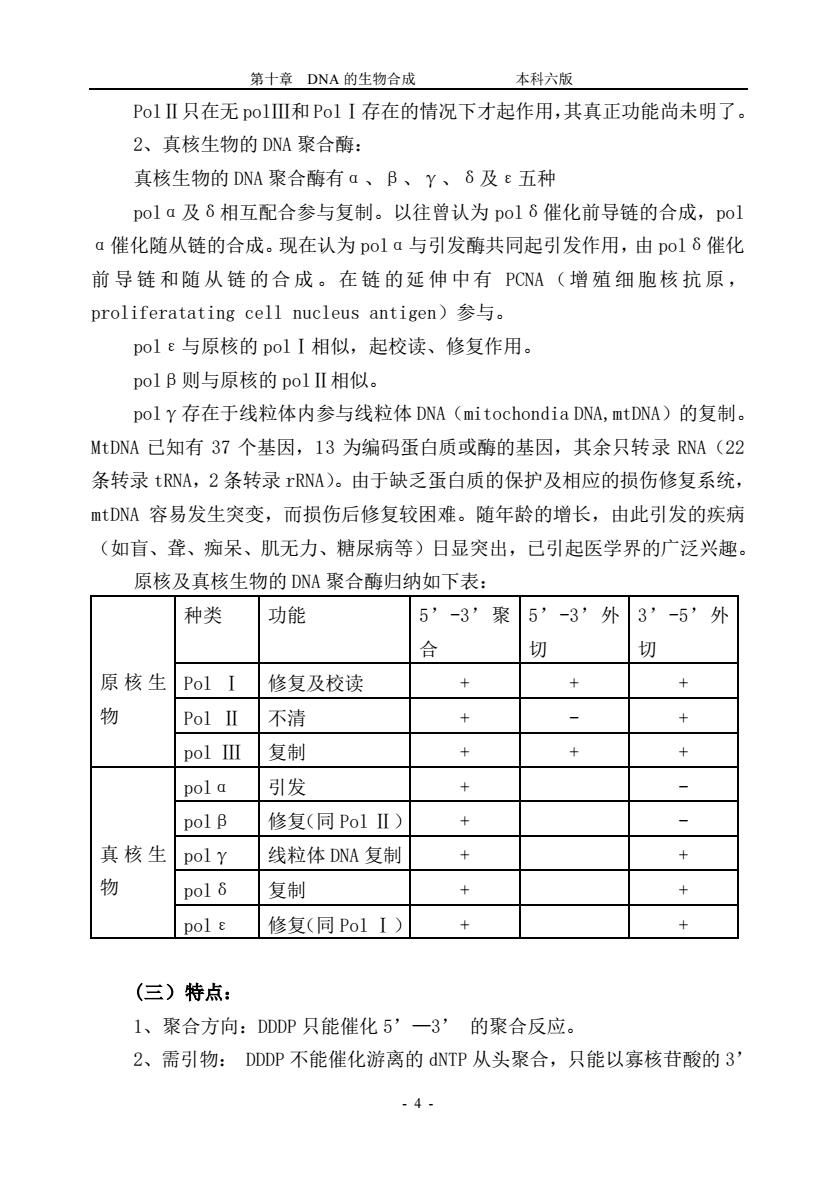
第十章DNA的生物合成本科六版PolII只在无polIⅢ和PolI存在的情况下才起作用,其真正功能尚未明了。2、真核生物的DNA聚合酶:真核生物的DNA聚合酶有α、β、、8及ε五种pola及8相互配合参与复制。以往曾认为pol催化前导链的合成,pola催化随从链的合成。现在认为pola与引发酶共同起引发作用,由pol8催化前导链和随从链的合成。在链的延伸中有PCNA(增殖细胞核抗原,proliferatatingcellnucleusantigen)参与。pol&与原核的polI相似,起校读、修复作用。polβ则与原核的polII相似。pol存在于线粒体内参与线粒体DNA(mitochondiaDNA,mtDNA)的复制。MtDNA已知有37个基因,13为编码蛋白质或酶的基因,其余只转录RNA(22条转录tRNA,2条转录rRNA)。由于缺乏蛋白质的保护及相应的损伤修复系统,mtDNA容易发生突变,而损伤后修复较困难。随年龄的增长,由此引发的疾病(如盲、聋、痴呆、肌无力、糖尿病等)日显突出,已引起医学界的广泛兴趣。原核及真核生物的DNA聚合酶归纳如下表:种类功能外3'-5'外5-3°聚5″-3*合切切原核生+Pol I修复及校读++物-不清++Pol II复制+pol III++引发+pol a一+pol β修复(同PolII)真核生++pol线粒体DNA复制物复制+pol 8+++pol e修复(同PolI)(三)特点:1、聚合方向:DDDP只能催化5一3的聚合反应。2、需引物:DDDP不能催化游离的dNTP从头聚合,只能以寡核苷酸的3-4
第十章 DNA 的生物合成 本科六版 - 4 - PolⅡ只在无 polⅢ和 PolⅠ存在的情况下才起作用,其真正功能尚未明了。 2、真核生物的 DNA 聚合酶: 真核生物的 DNA 聚合酶有α、β、γ、δ及ε五种 polα及δ相互配合参与复制。以往曾认为 polδ催化前导链的合成,pol α催化随从链的合成。现在认为 polα与引发酶共同起引发作用,由 polδ催化 前 导 链 和随 从 链 的合 成 。在 链 的延 伸 中有 PCNA (增 殖 细 胞核 抗 原 , proliferatating cell nucleus antigen)参与。 polε与原核的 polⅠ相似,起校读、修复作用。 polβ则与原核的 polⅡ相似。 polγ存在于线粒体内参与线粒体 DNA(mitochondia DNA,mtDNA)的复制。 MtDNA 已知有 37 个基因,13 为编码蛋白质或酶的基因,其余只转录 RNA(22 条转录 tRNA,2 条转录 rRNA)。由于缺乏蛋白质的保护及相应的损伤修复系统, mtDNA 容易发生突变,而损伤后修复较困难。随年龄的增长,由此引发的疾病 (如盲、聋、痴呆、肌无力、糖尿病等)日显突出,已引起医学界的广泛兴趣。 原核及真核生物的 DNA 聚合酶归纳如下表: 原核生 物 种类 功能 5’-3’聚 合 5’-3’外 切 3’-5’外 切 Pol Ⅰ 修复及校读 + + + Pol Ⅱ 不清 + - + pol Ⅲ 复制 + + + 真核生 物 polα 引发 + - polβ 修复(同 Pol Ⅱ) + - polγ 线粒体 DNA 复制 + + polδ 复制 + + polε 修复(同 Pol Ⅰ) + + (三)特点: 1、聚合方向:DDDP 只能催化 5’—3’ 的聚合反应。 2、需引物: DDDP 不能催化游离的 dNTP 从头聚合,只能以寡核苷酸的 3’

第十章DNA的生物合成本科六版一OH作为合成新DNA链的起点,该寡核苷酸称作引物。3、高度保真性:也就是对模板的高度依赖性(即与模板保持高度的一致性,体现在:1)DDDP能严格遵循碱基互补规律,选择与母链对应的dNTP与之配时,然后催化聚合反应。2)当复制出错时,DDDP有即时的校读功能(Proofread)二、解旋解链酶类:至少有三大类(一)解链酶(helicase):又名rep蛋白、DnaB蛋白1、作用:解开DNA双链,分开两链间的碱基对。2、特点:每解开一对碱基耗2个ATP3、种类:有DnaA、DnaB(即解链酶))、DnaC。DnaA辨认ori,DnaC辅助解链酶(DnaB)在起始点结合并结开双链。(二)DNA拓扑异构酶(拓扑酶:topoisomerase):拓扑是指物体或图象作弹性移位而又保持物体不变的性质。1、种类及作用形式:1)拓扑酶I只切断DNA双链中的一股,使螺旋松驰不打结,然后再将切口封起。2)拓扑酶II(即旋转酶,gyrase)可切断超螺旋DNA的两股链,切口处的两链旋转,使超螺旋结构松驰,在ATP存在时,DNA分子由松驰一一负超螺旋。最后将断端封好。2、作用:理顺DNA的结构,防止其打结、缠绕,配合复制过程。在DNA复制时,拓扑酶可松弛超螺旋结构,有利于复制叉的行进和DNA的合成;复制完成后,拓扑酶又可将DNA引入超螺旋结构,以形成染色质。(三)单链DNA结合蛋白(singlestrandedDNAbindingprotein,SSB)1、组成:SSB由177个氨基酸残基组成的相同四聚体,每个SSB可覆盖DNA单链上32个核苷酸。2、作用:1)与单链DNA结合,防止重新形成双螺旋2)保护单链DNA,免受Dnase作用而降解。- 5 -
第十章 DNA 的生物合成 本科六版 - 5 - —OH 作为合成新 DNA 链的起点,该寡核苷酸称作引物。 3、高度保真性:也就是对模板的高度依赖性(即与模板保持高度的一致 性),体现在: 1)DDDP 能严格遵循碱基互补规律,选择与母链对应的 dNTP 与之配时,然 后催化聚合反应。 2)当复制出错时,DDDP 有即时的校读功能(Proofread) 二、解旋解链酶类: 至少有三大类 (一)解链酶(helicase):又名 rep 蛋白、DnaB 蛋白 1、作用:解开 DNA 双链,分开两链间的碱基对。 2、特点:每解开一对碱基耗 2 个 ATP 3、种类:有 DnaA、DnaB(即解链酶))、DnaC。DnaA 辨认 ori,DnaC 辅助 解链酶(DnaB)在起始点结合并结开双链。 (二)DNA 拓扑异构酶(拓扑酶:topoisomerase): 拓扑是指物体或图象作弹性移位而又保持物体不变的性质。 1、种类及作用形式: 1)拓扑酶Ⅰ只切断 DNA 双链中的一股,使螺旋松驰不打结,然后再将切 口封起。 2)拓扑酶Ⅱ(即旋转酶,gyrase)可切断超螺旋 DNA 的两股链,切口处 的两链旋转,使超螺旋结构松驰,在 ATP 存在时,DNA 分子由松驰——负超螺 旋。最后将断端封好。 2、作用:理顺DNA的结构,防止其打结、缠绕,配合复制过程。 在DNA复制时,拓扑酶可松弛超螺旋结构,有利于复制叉的行进和DN A的合成;复制完成后,拓扑酶又可将DNA引入超螺旋结构,以形成染色质。 (三)单链 DNA 结合蛋白(single stranded DNA binding protein,SSB) 1、组成:SSB 由 177 个氨基酸残基组成的相同四聚体,每个 SSB 可覆盖 DNA单链上 32 个核苷酸。 2、作用:1)与单链 DNA 结合,防止重新形成双螺旋 2)保护单链 DNA,免受 Dnase 作用而降解