
复制:开始时,首先合成前体RNAIⅡ分子,在复制起始区上游 与DNA形成一种杂交体,杂交体被RnaseH酶所切割,从而释 放出3'-OH末端,作为供DNA聚合酶I合成DNA的引物。引导 质粒的复制过程。一旦DNA合成启动后,便不能改变其速度或 进程。 RNAI是由RNAII基因的反义链编码的1O8个碱基小分子转录 物。它折叠成可与新生RNA前体结合的三叶草结构,从而阻 止后者折叠成形成稳定杂交体所必需的二级结构。 另外,质粒本身编码的Rop蛋白能增强RNAI和RNAII的结合
复制:开始时,首先合成前体RNAⅡ分子,在复制起始区上游 与DNA形成一种杂交体,杂交体被RnaseH酶所切割,从而释 放出3′-OH末端,作为供DNA聚合酶Ⅰ合成DNA的引物。引导 质粒的复制过程。一旦DNA合成启动后,便不能改变其速度或 进程。 RNAⅠ是由RNAⅡ基因的反义链编码的108个碱基小分子转录 物。它折叠成可与新生RNA II前体结合的三叶草结构,从而阻 止后者折叠成形成稳定杂交体所必需的二级结构 。 另外,质粒本身编码的Rop蛋白能增强RNAⅠ和RNAⅡ的结合

因此,从本质上讲,CoE1质粒的复制启动显然是受一种负反馈 机理控制的。根据这种模型,细胞中RNAI分子的浓度是随着 质粒拷贝数的多寡而增减的。 例如,若细胞中质粒拷贝数下降到正常数值以下的水平, RNAI的浓度也就相应降低,于是质粒的复制也就受到较少的 抑制,结果导致其拷贝数的上升。 因此,只要削弱RNAI和RNAI之间的相互作用,将会增加质粒 的拷贝数
因此,从本质上讲,ColE1质粒的复制启动显然是受一种负反馈 机理控制的。根据这种模型,细胞中RNAⅠ分子的浓度是随着 质粒拷贝数的多寡而增减的。 例如,若细胞中质粒拷贝数下降到正常数值以下的水平, RNAⅠ的浓度也就相应降低,于是质粒的复制也就受到较少的 抑制,结果导致其拷贝数的上升。 因此,只要削弱RNAI和RNAII之间的相互作用,将会增加质粒 的拷贝数

②拷贝数 质粒拷贝数是指生长在标准的培养基条件下,每个细菌细胞 中所含有的质粒DNA分子的数目。 “严谨型”复制控制的质粒(stringent plasmid):低拷贝数 的质粒,每个宿主细胞中仅含有1-3拷贝; “松弛型”复制控制的质粒(relaxed plasmid:高拷贝数的 质粒,每个宿主细胞中可高达10-60拷贝
质粒拷贝数是指生长在标准的培养基条件下,每个细菌细胞 中所含有的质粒DNA分子的数目。 “严谨型”复制控制的质粒(stringent plasmid) :低拷贝数 的质粒,每个宿主细胞中仅含有1-3 拷贝; “松弛型”复制控制的质粒(relaxed plasmid) :高拷贝数的 质粒,每个宿主细胞中可高达10-60 拷贝。 ② 拷贝数
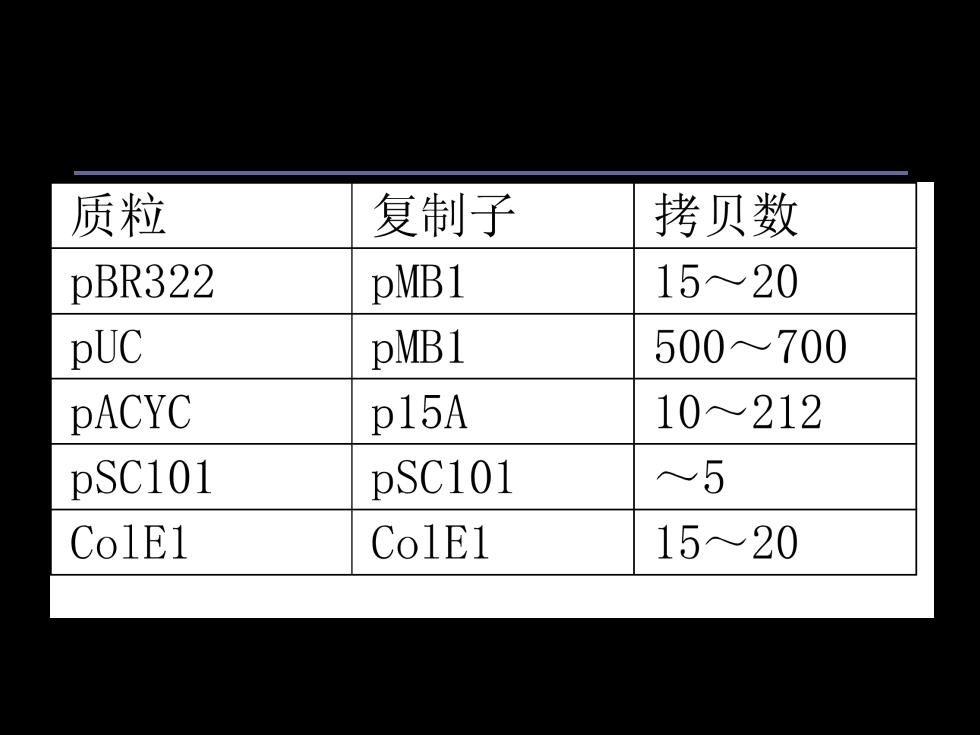
质粒 复制子 拷贝数 pBR322 pMB1 1520 pUC pMB1 500~700 PACYC p15A 10212 pSC101 pSC101 ~5 ColE1 ColE1 15~20

摇菌培养(pUC18)常加入氯霉素增加拷贝数 RNA分子作为调控物,不需要质粒编码的功能蛋白,而是完 全依靠宿主提供的半衰期较长的酶(DNA聚合酶I,DNA聚合 酶l,依赖于DNA的RNA聚合酶,以及宿主基因。 当存在抑制蛋白质合成并阻断细菌染色体复制的氯霉素或壮 观霉素等抗生素时,带有pMB1(或CoE1)复制子的质粒将继 续复制。 因此,即使蛋白质合成并非正在进行,复制依然能够我行我 素
摇菌培养(pUC18)常加入氯霉素增加拷贝数 RNA分子作为调控物,不需要质粒编码的功能蛋白,而是完 全依靠宿主提供的半衰期较长的酶(DNA聚合酶I,DNA聚合 酶III,依赖于DNA的RNA聚合酶,以及宿主基因。 当存在抑制蛋白质合成并阻断细菌染色体复制的氯霉素或壮 观霉素等抗生素时,带有pMB1(或ColE1)复制子的质粒将继 续复制。 因此,即使蛋白质合成并非正在进行,复制依然能够我行我 素