
1 浙江大学 遗传学第四章 1 第四章 连锁遗传和性连锁 浙江大学 遗传学第四章 2 本章重点 一、连锁遗传: 二对性状杂交有四种表现型,亲型多、重组型少; 杂种产生配子数不等,亲型相等、重组型相等。 二、连锁和交换机理: 粗线期交换、双线期交叉,非姐妹染色体交换。 三、交换值及其测定: 重组配子数/总配子数; 测交法测定,也可用 F2 材料进行估计。 浙江大学 遗传学第四章 3 本章重点 四、基因定位和连锁遗传图: 确定位置、距离,基因位于染色体上; 二点测验、三点测验; 连锁群、连锁遗传图。 五、性别决定: 与性别有关的一个或一对染色体称性染色体; 成对性染色体往往异型:XY型、ZW型; 性连锁。 浙江大学 遗传学第四章 4 1900年孟德尔遗传规律重新发现以后,生物界广泛 重视,进行了大量试验。 其中有些属于两对性状的遗传结果不符合独立分配 规律Î 摩尔根以果蝇为材料进行深入细致研究 Î 提出 连锁遗传规律Î 创立基因论 Î 认为基因成直线排列在 染色体上,进一步发展为细胞遗传学。 浙江大学 遗传学第四章 5 第一节 连锁与交换 浙江大学 遗传学第四章 6 一、连 锁
1 浙江大学 遗传学第四章 1 第四章 连锁遗传和性连锁 浙江大学 遗传学第四章 2 本章重点 一、连锁遗传: 二对性状杂交有四种表现型,亲型多、重组型少; 杂种产生配子数不等,亲型相等、重组型相等。 二、连锁和交换机理: 粗线期交换、双线期交叉,非姐妹染色体交换。 三、交换值及其测定: 重组配子数/总配子数; 测交法测定,也可用 F2 材料进行估计。 浙江大学 遗传学第四章 3 本章重点 四、基因定位和连锁遗传图: 确定位置、距离,基因位于染色体上; 二点测验、三点测验; 连锁群、连锁遗传图。 五、性别决定: 与性别有关的一个或一对染色体称性染色体; 成对性染色体往往异型:XY型、ZW型; 性连锁。 浙江大学 遗传学第四章 4 1900年孟德尔遗传规律重新发现以后,生物界广泛 重视,进行了大量试验。 其中有些属于两对性状的遗传结果不符合独立分配 规律Î 摩尔根以果蝇为材料进行深入细致研究 Î 提出 连锁遗传规律Î 创立基因论 Î 认为基因成直线排列在 染色体上,进一步发展为细胞遗传学。 浙江大学 遗传学第四章 5 第一节 连锁与交换 浙江大学 遗传学第四章 6 一、连 锁
2 浙江大学 遗传学第四章 7 (一)、性状连锁遗传的发现 浙江大学 遗传学第四章 8 1906年,贝特生(Bateson W.) 和贝拉特(Punnett R. C.)在香豌豆 的二对性状杂交试验中 ¨ 首先发现 性状连锁遗传现象。 贝特生(1861~1926): 英国生物学家, 曾经重复过孟德尔的实验 浙江大学 遗传学第四章 9 P 紫花、长花粉粒(PPLL) × 红花、圆花粉粒(ppll) ↓ F1 紫、长 PpLl ↓⊗ F2 紫、长 紫、圆 红、长 红、圆 P_L_ P_ll ppL_ ppll 总数 实际个体数 4831 390 393 1338 6952 按9:3:3:1推算 3910.5 1303.5 1303.5 434.5 6952 以上结果表明F2: ①. 同样出现四种表现型; ②. 不符合9:3:3:1; ③. 亲本组合数偏多,重新组合数偏少(与理论数相比)。 第一个试验: 浙江大学 遗传学第四章 10 P 紫花、圆花粉粒(PPll)× 红花、长花粉粒(ppLL) ↓ F1 紫、长PpLl ↓⊗ F2 紫、长 紫、圆 红、长 红、圆 P_L_ P_ll ppL_ ppll 总数 实际个体数 226 95 97 1 419 按9:3:3:1推算 235.8 78.5 78.5 26.2 419 结果与第一个试验情况相同。 第二个试验: 浙江大学 遗传学第四章 11 连锁遗传:原来亲本所具有的两个性状,在F2连系在一起 遗传的现象。 相引组:甲乙两个显性性状,连系在一起遗传、而甲乙两 个隐性性状连系在一起的杂交组合。 如:PL/pl。 相斥组:甲显性性状和乙隐性性状连系在一起遗传,而乙 显性性状和甲隐性性状连系在一起的杂交组合。 如:Pl/pL。 浙江大学 遗传学第四章 12 (二)、连锁遗传的解释
2 浙江大学 遗传学第四章 7 (一)、性状连锁遗传的发现 浙江大学 遗传学第四章 8 1906年,贝特生(Bateson W.) 和贝拉特(Punnett R. C.)在香豌豆 的二对性状杂交试验中 ¨ 首先发现 性状连锁遗传现象。 贝特生(1861~1926): 英国生物学家, 曾经重复过孟德尔的实验 浙江大学 遗传学第四章 9 P 紫花、长花粉粒(PPLL) × 红花、圆花粉粒(ppll) ↓ F1 紫、长 PpLl ↓⊗ F2 紫、长 紫、圆 红、长 红、圆 P_L_ P_ll ppL_ ppll 总数 实际个体数 4831 390 393 1338 6952 按9:3:3:1推算 3910.5 1303.5 1303.5 434.5 6952 以上结果表明F2: ①. 同样出现四种表现型; ②. 不符合9:3:3:1; ③. 亲本组合数偏多,重新组合数偏少(与理论数相比)。 第一个试验: 浙江大学 遗传学第四章 10 P 紫花、圆花粉粒(PPll)× 红花、长花粉粒(ppLL) ↓ F1 紫、长PpLl ↓⊗ F2 紫、长 紫、圆 红、长 红、圆 P_L_ P_ll ppL_ ppll 总数 实际个体数 226 95 97 1 419 按9:3:3:1推算 235.8 78.5 78.5 26.2 419 结果与第一个试验情况相同。 第二个试验: 浙江大学 遗传学第四章 11 连锁遗传:原来亲本所具有的两个性状,在F2连系在一起 遗传的现象。 相引组:甲乙两个显性性状,连系在一起遗传、而甲乙两 个隐性性状连系在一起的杂交组合。 如:PL/pl。 相斥组:甲显性性状和乙隐性性状连系在一起遗传,而乙 显性性状和甲隐性性状连系在一起的杂交组合。 如:Pl/pL。 浙江大学 遗传学第四章 12 (二)、连锁遗传的解释

3 浙江大学 遗传学第四章 13 试验结果是否受分离规律支配? 第一个试验: 紫花:红花 (4831+390):(1338+393)=5221:1731 ≈ 3:1 长花粉:短花粉 (4831+393):(1338+390)=5224:1728 ≈ 3:1 第二个试验: 紫花:红花 (226+95):(97+1)=321:98 ≈ 3:1 长花粉:短花粉 (226+97):(95+1)=323:96 ≈ 3:1 } 以上结果都受分离规律支配,但不符合独立分配规律。 } F2不符合9:3:3:1,则说明F1产生的四种配子不等。 可用测交法加以验证,∵测交后代的表现型种类以及 比例可反映出F1配子的种类和比例。 浙江大学 遗传学第四章 14 利用测交法验证连锁遗传现象: 特点:连锁遗传的表现为: ♥ 两个亲本型配子数是相等,>50%; ♥ 两个重组型配子数相等,<50%。 1.相引组: 玉米(种子性状当代即可观察): 有色、饱满CCShSh × 无色、凹陷ccshsh ↓ F1 有色饱满 × 无色凹陷 CcShsh ccshsh 配子 CSh Csh cSh csh csh Ft CcShsh Ccshsh ccShsh ccshsh 有色饱满 有色凹陷 无色饱满 无色凹陷 总数 粒数 4032 149 152 4035 8368 % 48.2 1.8 1.8 48.2 上述结果证实F1产生的四种配子不等: 不是1:1:1:1 (25:25:25:25%)。其中(Ft) 亲本组合 = (4032+4035)/8368×100% = 96.4% 重新组合 = (149+152)/8368×100% = 3.6% 摩尔根(Morgan T. H.)等以果蝇为材料进行测交的结果: 红眼长翅 pr+pr+vg+vg+ × 紫眼正常翅 prprvgvg ↓ F1 红眼长翅 × 紫眼正常翅 pr+prvg+vg prprvgvg 配子 pr+vg+ prvg+ pr+vg prvg prvg Ft pr+prvg+vg prprvg+vg pr+prvgvg prprvgvg 红眼长翅 紫眼长翅 红眼正常翅 紫眼正常翅 总数 个数 1339 154 151 1195 2839 结果:亲本组合=((1339+1195)/2839)×100%=89.26% 重新组合=((154+151)/2839)×100%=10.74% ∴ 证实F1所成的四种配子数不等,两种亲型配子多,两种重组型少,分别 接近1:1。 浙江大学 遗传学第四章 17 亲本具有的两对非等位基因(Cc和Shsh 或 pr+pr 和vg+vg)不是独立分配,而是连系在一起遗传,如 C-Sh、c-sh或pr+vg+和prvg常常连系在一起。 ∴ F1配子中总是亲本型配子(CSh和csh或pr+vg+ 和prvg)数偏多,重新组合配子(Csh、cSh或pr+vg 和prvg+)数偏少。 2.相斥组: 玉米: 有色、凹陷CCshsh × 无色、饱满ccShSh ↓ F1 有色饱满 × 无色凹陷 CcShsh ccshsh 配子 CSh Csh cSh csh csh Ft CcShsh Ccshsh ccShsh ccshsh 有色饱满 有色凹陷 无色饱满 无色凹陷 总数 粒数 638 21379 21096 672 43785 % 1.5 48.5 48.5 1.5 结果:亲本组合=(21379+21096)/43785×100%=97.01% 重新组合=(638+672)/843785×100%=2.99% ∴ 相斥组的结果与相引组结果一致,同样证实F1所成的四种配子数 不等,C-sh、c-Sh连系在一起的配子为多
3 浙江大学 遗传学第四章 13 试验结果是否受分离规律支配? 第一个试验: 紫花:红花 (4831+390):(1338+393)=5221:1731 ≈ 3:1 长花粉:短花粉 (4831+393):(1338+390)=5224:1728 ≈ 3:1 第二个试验: 紫花:红花 (226+95):(97+1)=321:98 ≈ 3:1 长花粉:短花粉 (226+97):(95+1)=323:96 ≈ 3:1 } 以上结果都受分离规律支配,但不符合独立分配规律。 } F2不符合9:3:3:1,则说明F1产生的四种配子不等。 可用测交法加以验证,∵测交后代的表现型种类以及 比例可反映出F1配子的种类和比例。 浙江大学 遗传学第四章 14 利用测交法验证连锁遗传现象: 特点:连锁遗传的表现为: ♥ 两个亲本型配子数是相等,>50%; ♥ 两个重组型配子数相等,<50%。 1.相引组: 玉米(种子性状当代即可观察): 有色、饱满CCShSh × 无色、凹陷ccshsh ↓ F1 有色饱满 × 无色凹陷 CcShsh ccshsh 配子 CSh Csh cSh csh csh Ft CcShsh Ccshsh ccShsh ccshsh 有色饱满 有色凹陷 无色饱满 无色凹陷 总数 粒数 4032 149 152 4035 8368 % 48.2 1.8 1.8 48.2 上述结果证实F1产生的四种配子不等: 不是1:1:1:1 (25:25:25:25%)。其中(Ft) 亲本组合 = (4032+4035)/8368×100% = 96.4% 重新组合 = (149+152)/8368×100% = 3.6% 摩尔根(Morgan T. H.)等以果蝇为材料进行测交的结果: 红眼长翅 pr+pr+vg+vg+ × 紫眼正常翅 prprvgvg ↓ F1 红眼长翅 × 紫眼正常翅 pr+prvg+vg prprvgvg 配子 pr+vg+ prvg+ pr+vg prvg prvg Ft pr+prvg+vg prprvg+vg pr+prvgvg prprvgvg 红眼长翅 紫眼长翅 红眼正常翅 紫眼正常翅 总数 个数 1339 154 151 1195 2839 结果:亲本组合=((1339+1195)/2839)×100%=89.26% 重新组合=((154+151)/2839)×100%=10.74% ∴ 证实F1所成的四种配子数不等,两种亲型配子多,两种重组型少,分别 接近1:1。 浙江大学 遗传学第四章 17 亲本具有的两对非等位基因(Cc和Shsh 或 pr+pr 和vg+vg)不是独立分配,而是连系在一起遗传,如 C-Sh、c-sh或pr+vg+和prvg常常连系在一起。 ∴ F1配子中总是亲本型配子(CSh和csh或pr+vg+ 和prvg)数偏多,重新组合配子(Csh、cSh或pr+vg 和prvg+)数偏少。 2.相斥组: 玉米: 有色、凹陷CCshsh × 无色、饱满ccShSh ↓ F1 有色饱满 × 无色凹陷 CcShsh ccshsh 配子 CSh Csh cSh csh csh Ft CcShsh Ccshsh ccShsh ccshsh 有色饱满 有色凹陷 无色饱满 无色凹陷 总数 粒数 638 21379 21096 672 43785 % 1.5 48.5 48.5 1.5 结果:亲本组合=(21379+21096)/43785×100%=97.01% 重新组合=(638+672)/843785×100%=2.99% ∴ 相斥组的结果与相引组结果一致,同样证实F1所成的四种配子数 不等,C-sh、c-Sh连系在一起的配子为多
4 浙江大学 遗传学第四章 19 果蝇(相斥组): 红眼正常翅pr+pr+vgvg × 紫眼长翅prprvg+vg+ ↓ F1 红眼长翅 × 紫眼正常翅 pr+prvg+vg prprvgvg 配子 pr+vg+ prvg+ pr+vg prvg prvg Ft pr+prvg+vg prprvg+vg pr+prvgvg prprvgvg 红眼长翅 紫眼长翅 红眼正常翅 紫眼正常翅 总数 个数 157 1067 965 146 2335 结果:亲本组合=((1067+965)/2335)×100%=87.02% 重新组合=((157+146)/2335)×100%=12.98% ∴ 证实F1所成的四种配子数不等。 浙江大学 遗传学第四章 20 ♥上述结果均说明重组型配子数占总配子数的百分率< 50%。 重组率(交换值):重组型的配子百分数称为重组率。 ♥ 当两对基因为连锁遗传时,其重组率总是<50。 ∵相引组或相斥组中一对同源染色体的四条非姐妹染色单 体,两个基因之间的染色体区段内 仅有两条非姐妹染色单体发生交换 (crossing over)。 ∴重组型配子数目只是少数。 浙江大学 遗传学第四章 21 (三)、完全连锁与不完全连锁: 浙江大学 遗传学第四章 22 生物性状很多,控制这些性状的基因自然也多,而生物 的染色体数目有限 ¨ 必有许多基因位于同一染色体上 ¨ 引起连锁遗传。 ♣ 连锁:若干非等位基因位于同一染色体而发生连系遗传 的现象。 ♣ 完全连锁:同源染色体上非等位基因间不能发生非姐妹 染色单体之间的交换 ¨ F1只产生两种亲型配子、其自交或 测交后代个体的表现型均为亲本组合。 AB ab P AB × ab AB F1 ab ⊗ × ab ab ♂ ♂ ♀ AB ab ♀ ab AB AB AB AB AB AB ab ab ab AB ab ab ab ab ab ab 完全连锁 3:1 测交 1:1 浙江大学 遗传学第四章 24 如1910年摩尔根在果蝇试验中也发现: P b vg+ b+ vg 黑身长翅 b vg+ × b+ vg 灰身短翅 b vg+ Ft 灰身长翅 b+ vg b+ vg 近亲繁殖 测交 × b+ vg ♂ ♂ ♀ b vg+ b+ vg ♀ b+ vg b vg+ b vg+ b vg+ b vg+ b vg+ b vg+ b+ vg b+ vg b+ vg b vg+ b+ vg b+ vg b+ vg b+ vg b+ vg b+ vg 灰身长翅:黑身短翅 = 3:1 灰身长翅:黑身短翅 = 1:1
4 浙江大学 遗传学第四章 19 果蝇(相斥组): 红眼正常翅pr+pr+vgvg × 紫眼长翅prprvg+vg+ ↓ F1 红眼长翅 × 紫眼正常翅 pr+prvg+vg prprvgvg 配子 pr+vg+ prvg+ pr+vg prvg prvg Ft pr+prvg+vg prprvg+vg pr+prvgvg prprvgvg 红眼长翅 紫眼长翅 红眼正常翅 紫眼正常翅 总数 个数 157 1067 965 146 2335 结果:亲本组合=((1067+965)/2335)×100%=87.02% 重新组合=((157+146)/2335)×100%=12.98% ∴ 证实F1所成的四种配子数不等。 浙江大学 遗传学第四章 20 ♥上述结果均说明重组型配子数占总配子数的百分率< 50%。 重组率(交换值):重组型的配子百分数称为重组率。 ♥ 当两对基因为连锁遗传时,其重组率总是<50。 ∵相引组或相斥组中一对同源染色体的四条非姐妹染色单 体,两个基因之间的染色体区段内 仅有两条非姐妹染色单体发生交换 (crossing over)。 ∴重组型配子数目只是少数。 浙江大学 遗传学第四章 21 (三)、完全连锁与不完全连锁: 浙江大学 遗传学第四章 22 生物性状很多,控制这些性状的基因自然也多,而生物 的染色体数目有限 ¨ 必有许多基因位于同一染色体上 ¨ 引起连锁遗传。 ♣ 连锁:若干非等位基因位于同一染色体而发生连系遗传 的现象。 ♣ 完全连锁:同源染色体上非等位基因间不能发生非姐妹 染色单体之间的交换 ¨ F1只产生两种亲型配子、其自交或 测交后代个体的表现型均为亲本组合。 AB ab P AB × ab AB F1 ab ⊗ × ab ab ♂ ♂ ♀ AB ab ♀ ab AB AB AB AB AB AB ab ab ab AB ab ab ab ab ab ab 完全连锁 3:1 测交 1:1 浙江大学 遗传学第四章 24 如1910年摩尔根在果蝇试验中也发现: P b vg+ b+ vg 黑身长翅 b vg+ × b+ vg 灰身短翅 b vg+ Ft 灰身长翅 b+ vg b+ vg 近亲繁殖 测交 × b+ vg ♂ ♂ ♀ b vg+ b+ vg ♀ b+ vg b vg+ b vg+ b vg+ b vg+ b vg+ b vg+ b+ vg b+ vg b+ vg b vg+ b+ vg b+ vg b+ vg b+ vg b+ vg b+ vg 灰身长翅:黑身短翅 = 3:1 灰身长翅:黑身短翅 = 1:1
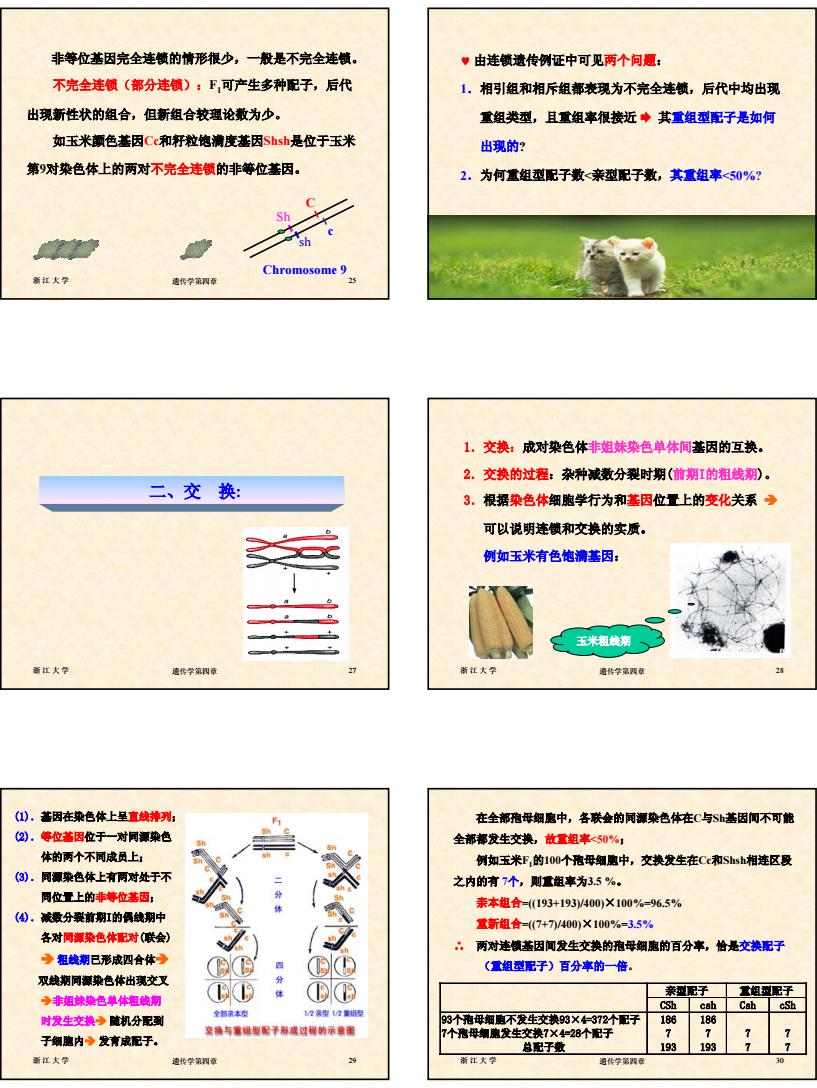
5 浙江大学 遗传学第四章 25 非等位基因完全连锁的情形很少,一般是不完全连锁。 不完全连锁(部分连锁):F1可产生多种配子,后代 出现新性状的组合,但新组合较理论数为少。 如玉米颜色基因Cc和籽粒饱满度基因Shsh是位于玉米 第9对染色体上的两对不完全连锁的非等位基因。 C c Sh sh Chromosome 9 浙江大学 遗传学第四章 26 ♥ 由连锁遗传例证中可见两个问题: 1.相引组和相斥组都表现为不完全连锁,后代中均出现 重组类型,且重组率很接近 ¨ 其重组型配子是如何 出现的? 2.为何重组型配子数<亲型配子数,其重组率<50%? 浙江大学 遗传学第四章 27 二、交 换: 浙江大学 遗传学第四章 28 1.交换:成对染色体非姐妹染色单体间基因的互换。 2.交换的过程:杂种减数分裂时期(前期I的粗线期)。 3.根据染色体细胞学行为和基因位置上的变化关系 Î 可以说明连锁和交换的实质。 例如玉米有色饱满基因: 玉米粗线期 浙江大学 遗传学第四章 29 (1).基因在染色体上呈直线排列; (2).等位基因位于一对同源染色 体的两个不同成员上; (3).同源染色体上有两对处于不 同位置上的非等位基因; (4).减数分裂前期I的偶线期中 各对同源染色体配对(联会) Î 粗线期已形成四合体Î 双线期同源染色体出现交叉 Î非姐妹染色单体粗线期 时发生交换Î 随机分配到 子细胞内Î 发育成配子。 浙江大学 遗传学第四章 30 在全部孢母细胞中,各联会的同源染色体在C与Sh基因间不可能 全部都发生交换,故重组率<50%; 例如玉米F1的100个孢母细胞中,交换发生在Cc和Shsh相连区段 之内的有 7个,则重组率为3.5 %。 亲本组合=((193+193)/400)×100%=96.5% 重新组合=((7+7)/400)×100%=3.5% ∴ 两对连锁基因间发生交换的孢母细胞的百分率,恰是交换配子 (重组型配子)百分率的一倍。 亲型配子 重组型配子 CSh csh Csh cSh 93个孢母细胞不发生交换93×4=372个配子 186 186 7个孢母细胞发生交换7×4=28个配子 7 7 7 7 总配子数 193 193 7 7
5 浙江大学 遗传学第四章 25 非等位基因完全连锁的情形很少,一般是不完全连锁。 不完全连锁(部分连锁):F1可产生多种配子,后代 出现新性状的组合,但新组合较理论数为少。 如玉米颜色基因Cc和籽粒饱满度基因Shsh是位于玉米 第9对染色体上的两对不完全连锁的非等位基因。 C c Sh sh Chromosome 9 浙江大学 遗传学第四章 26 ♥ 由连锁遗传例证中可见两个问题: 1.相引组和相斥组都表现为不完全连锁,后代中均出现 重组类型,且重组率很接近 ¨ 其重组型配子是如何 出现的? 2.为何重组型配子数<亲型配子数,其重组率<50%? 浙江大学 遗传学第四章 27 二、交 换: 浙江大学 遗传学第四章 28 1.交换:成对染色体非姐妹染色单体间基因的互换。 2.交换的过程:杂种减数分裂时期(前期I的粗线期)。 3.根据染色体细胞学行为和基因位置上的变化关系 Î 可以说明连锁和交换的实质。 例如玉米有色饱满基因: 玉米粗线期 浙江大学 遗传学第四章 29 (1).基因在染色体上呈直线排列; (2).等位基因位于一对同源染色 体的两个不同成员上; (3).同源染色体上有两对处于不 同位置上的非等位基因; (4).减数分裂前期I的偶线期中 各对同源染色体配对(联会) Î 粗线期已形成四合体Î 双线期同源染色体出现交叉 Î非姐妹染色单体粗线期 时发生交换Î 随机分配到 子细胞内Î 发育成配子。 浙江大学 遗传学第四章 30 在全部孢母细胞中,各联会的同源染色体在C与Sh基因间不可能 全部都发生交换,故重组率<50%; 例如玉米F1的100个孢母细胞中,交换发生在Cc和Shsh相连区段 之内的有 7个,则重组率为3.5 %。 亲本组合=((193+193)/400)×100%=96.5% 重新组合=((7+7)/400)×100%=3.5% ∴ 两对连锁基因间发生交换的孢母细胞的百分率,恰是交换配子 (重组型配子)百分率的一倍。 亲型配子 重组型配子 CSh csh Csh cSh 93个孢母细胞不发生交换93×4=372个配子 186 186 7个孢母细胞发生交换7×4=28个配子 7 7 7 7 总配子数 193 193 7 7