1 第八章 细胞质遗传 浙江大学 遗传学第八章 2 1. 细胞质遗传与核遗传的差异。 2. 细胞质遗传的特点。 3. 质核基因间的关系。 4. 雄性不育: ①.核不育型、质核互作不育型; ②.孢子体不育、配子体不育; ③.生产应用。 本章重点 第一节 细胞质遗传的特点 浙江大学 遗传学第八章 4 一、细胞质遗传的概念: 遗 传 物 质 核基因组 染色体 细胞质 基因组 细胞器 基因组 非细胞器 基因组 叶绿体基因组(ctDNA) 线粒体基因组(mtDNA) 中心粒基因组(centro DNA) 动粒基因组(kinto DNA) 膜体系基因组(membrane DNA) 共生体基因组(symbiont DNA) 细菌质粒基因组(plasmid DNA) 基因(核DNA) 浙江大学 遗传学第八章 5 细胞质遗传: 由胞质遗传物质引起的遗传现象(又称非染色体遗传、 非孟德尔遗传、染色体外遗传、核外遗传、母性遗传)。 细胞质基因组: 所有细胞器和细胞质颗粒中遗传物质 的统称。 浙江大学 遗传学第八章 6 1.特点: ①.正交和反交的遗传表现 不同。 核遗传:表现相同,其 遗传物质由雌核和雄核共同 提供; 质遗传:表现不同,某些 性状表现于母本时才能遗传 给子代,故又称母性遗传。 二、细胞质遗传的特点:
1 第八章 细胞质遗传 浙江大学 遗传学第八章 2 1. 细胞质遗传与核遗传的差异。 2. 细胞质遗传的特点。 3. 质核基因间的关系。 4. 雄性不育: ①.核不育型、质核互作不育型; ②.孢子体不育、配子体不育; ③.生产应用。 本章重点 第一节 细胞质遗传的特点 浙江大学 遗传学第八章 4 一、细胞质遗传的概念: 遗 传 物 质 核基因组 染色体 细胞质 基因组 细胞器 基因组 非细胞器 基因组 叶绿体基因组(ctDNA) 线粒体基因组(mtDNA) 中心粒基因组(centro DNA) 动粒基因组(kinto DNA) 膜体系基因组(membrane DNA) 共生体基因组(symbiont DNA) 细菌质粒基因组(plasmid DNA) 基因(核DNA) 浙江大学 遗传学第八章 5 细胞质遗传: 由胞质遗传物质引起的遗传现象(又称非染色体遗传、 非孟德尔遗传、染色体外遗传、核外遗传、母性遗传)。 细胞质基因组: 所有细胞器和细胞质颗粒中遗传物质 的统称。 浙江大学 遗传学第八章 6 1.特点: ①.正交和反交的遗传表现 不同。 核遗传:表现相同,其 遗传物质由雌核和雄核共同 提供; 质遗传:表现不同,某些 性状表现于母本时才能遗传 给子代,故又称母性遗传。 二、细胞质遗传的特点:

2 浙江大学 遗传学第八章 7 ②.连续回交,母本核基因可被全部 置换掉,但由母本细胞质基因所 控制的性状仍不会消失; ③.由细胞质中的附加体或共生体 决定的性状,其表现往往类似 病毒的转导或感染,即可传递 给其它细胞。 ④.基因定位困难: 遗传方式是非孟德尔遗传, 杂交后代不表现有比例的分离。 带有胞质基因的细胞器在 细胞分裂时分配是不均匀的。 浙江大学 遗传学第八章 8 2.母性遗传: 真核生物有性过程: 卵细胞:有细胞核、大量的细胞质和细胞器。 ∴能为子代提供核基因和全部或绝大部分胞质基因。 精细胞:只有细胞核,细胞质或细胞器极少或没有。 ∴只能提供其核基因,不能或极少提供胞质基因。 ∴一切受细胞质基因所决定的性状,其遗传信息一般只能 通过卵细胞传给子代,而不能通过精细胞遗传给子代。 卵细胞 精细胞 第二节 母性影响 浙江大学 遗传学第八章 10 一、母性影响的概念: 母性影响:由核基因的产物积累在卵细胞中的物质 所引起的一种遗传现象。 ∴母性影响不属于胞质遗传的范畴。 二、母性影响的特点: 下一代表现型受上一代母体基因的影响。 浙江大学 遗传学第八章 11 三、实例: 椎实螺的外壳旋转方向的遗传。 椎实螺是一种♀、♂同体的软体 动物,每一个体又能同时产生卵子和 精子,但一般通过异体受精进行繁殖。 ∴椎实螺即可进行异体杂交、又可单独进行个体的自体 受精。 椎实螺外壳的旋转方向有左旋和右旋之分,属于一对 相对性状。 浙江大学 遗传学第八章 12 3.杂交试验: ①.过程:椎实螺进行正反交,F1旋转方向都与各自母本相似, 即右旋或左旋,F2均为右旋,F3才出现右旋和左旋的 分离
2 浙江大学 遗传学第八章 7 ②.连续回交,母本核基因可被全部 置换掉,但由母本细胞质基因所 控制的性状仍不会消失; ③.由细胞质中的附加体或共生体 决定的性状,其表现往往类似 病毒的转导或感染,即可传递 给其它细胞。 ④.基因定位困难: 遗传方式是非孟德尔遗传, 杂交后代不表现有比例的分离。 带有胞质基因的细胞器在 细胞分裂时分配是不均匀的。 浙江大学 遗传学第八章 8 2.母性遗传: 真核生物有性过程: 卵细胞:有细胞核、大量的细胞质和细胞器。 ∴能为子代提供核基因和全部或绝大部分胞质基因。 精细胞:只有细胞核,细胞质或细胞器极少或没有。 ∴只能提供其核基因,不能或极少提供胞质基因。 ∴一切受细胞质基因所决定的性状,其遗传信息一般只能 通过卵细胞传给子代,而不能通过精细胞遗传给子代。 卵细胞 精细胞 第二节 母性影响 浙江大学 遗传学第八章 10 一、母性影响的概念: 母性影响:由核基因的产物积累在卵细胞中的物质 所引起的一种遗传现象。 ∴母性影响不属于胞质遗传的范畴。 二、母性影响的特点: 下一代表现型受上一代母体基因的影响。 浙江大学 遗传学第八章 11 三、实例: 椎实螺的外壳旋转方向的遗传。 椎实螺是一种♀、♂同体的软体 动物,每一个体又能同时产生卵子和 精子,但一般通过异体受精进行繁殖。 ∴椎实螺即可进行异体杂交、又可单独进行个体的自体 受精。 椎实螺外壳的旋转方向有左旋和右旋之分,属于一对 相对性状。 浙江大学 遗传学第八章 12 3.杂交试验: ①.过程:椎实螺进行正反交,F1旋转方向都与各自母本相似, 即右旋或左旋,F2均为右旋,F3才出现右旋和左旋的 分离

3 浙江大学 遗传学第八章 13 正交 反交 ♀(右旋)SS×♂(左旋)ss ♀(左旋)ss×♂(右旋)SS ↓异体受精 ↓异体受精 F1 全部为右旋(Ss) 全为左旋(Ss) ↓自体受精 ↓自体受精 F2 1SS∶2Ss∶1ss(均右旋) 1SS∶2Ss∶1ss(均右旋) ↓ ↓ ↓ ↓ ↓ ↓ F3 右旋 右旋 左旋 右旋 右旋 左旋 (SS)(分离)(ss) (SS)(分离)(ss) 3 ∶ 1 3 ∶ 1 ∴ 如果试验只进行到F1,很可能被误认为细胞质遗传。 ∵ F1的旋转方向同母本一致。 浙江大学 遗传学第八章 14 ②. 原因:椎实螺外壳旋转方向是由受精卵分裂时纺锤体分裂 方向决定的,并由受精前的母体基因型决定。 右旋──受精卵纺锤体向中线右侧分裂; 左旋──受精卵纺锤体向中线左侧分裂。 ∴母体基因型 受精卵纺锤体分裂方向 椎实螺 外壳旋转方向。 浙江大学 遗传学第八章 15 ③.前定作用或延迟遗传: 由受精前母体卵细胞基因型决定子代性状表现的 母性影响。 第三节 叶绿体遗传 浙江大学 遗传学第八章 17 一、叶绿体遗传的表现: 1.紫茉莉花斑性状遗传: ①.紫茉莉花斑植株: 着生绿色、白色和 花斑三种枝条,且白色 和绿色组织间有明显的 界限。 ①.柯伦斯杂交试验: 接受花粉的枝条 提供花粉的枝条 杂种植株的表现 白色 白色、绿色、花斑 均为白色 绿色 白色、绿色、花斑 均为绿色 花斑 白色、绿色、花斑 白色、绿色、花斑 (但不成比例) 杂种植株所表现的性状完全由母本枝条所决定,与提供花粉 的父本枝条无关。 ∴控制紫茉莉花斑性状的遗传物质是通过母本传递的
3 浙江大学 遗传学第八章 13 正交 反交 ♀(右旋)SS×♂(左旋)ss ♀(左旋)ss×♂(右旋)SS ↓异体受精 ↓异体受精 F1 全部为右旋(Ss) 全为左旋(Ss) ↓自体受精 ↓自体受精 F2 1SS∶2Ss∶1ss(均右旋) 1SS∶2Ss∶1ss(均右旋) ↓ ↓ ↓ ↓ ↓ ↓ F3 右旋 右旋 左旋 右旋 右旋 左旋 (SS)(分离)(ss) (SS)(分离)(ss) 3 ∶ 1 3 ∶ 1 ∴ 如果试验只进行到F1,很可能被误认为细胞质遗传。 ∵ F1的旋转方向同母本一致。 浙江大学 遗传学第八章 14 ②. 原因:椎实螺外壳旋转方向是由受精卵分裂时纺锤体分裂 方向决定的,并由受精前的母体基因型决定。 右旋──受精卵纺锤体向中线右侧分裂; 左旋──受精卵纺锤体向中线左侧分裂。 ∴母体基因型 受精卵纺锤体分裂方向 椎实螺 外壳旋转方向。 浙江大学 遗传学第八章 15 ③.前定作用或延迟遗传: 由受精前母体卵细胞基因型决定子代性状表现的 母性影响。 第三节 叶绿体遗传 浙江大学 遗传学第八章 17 一、叶绿体遗传的表现: 1.紫茉莉花斑性状遗传: ①.紫茉莉花斑植株: 着生绿色、白色和 花斑三种枝条,且白色 和绿色组织间有明显的 界限。 ①.柯伦斯杂交试验: 接受花粉的枝条 提供花粉的枝条 杂种植株的表现 白色 白色、绿色、花斑 均为白色 绿色 白色、绿色、花斑 均为绿色 花斑 白色、绿色、花斑 白色、绿色、花斑 (但不成比例) 杂种植株所表现的性状完全由母本枝条所决定,与提供花粉 的父本枝条无关。 ∴控制紫茉莉花斑性状的遗传物质是通过母本传递的
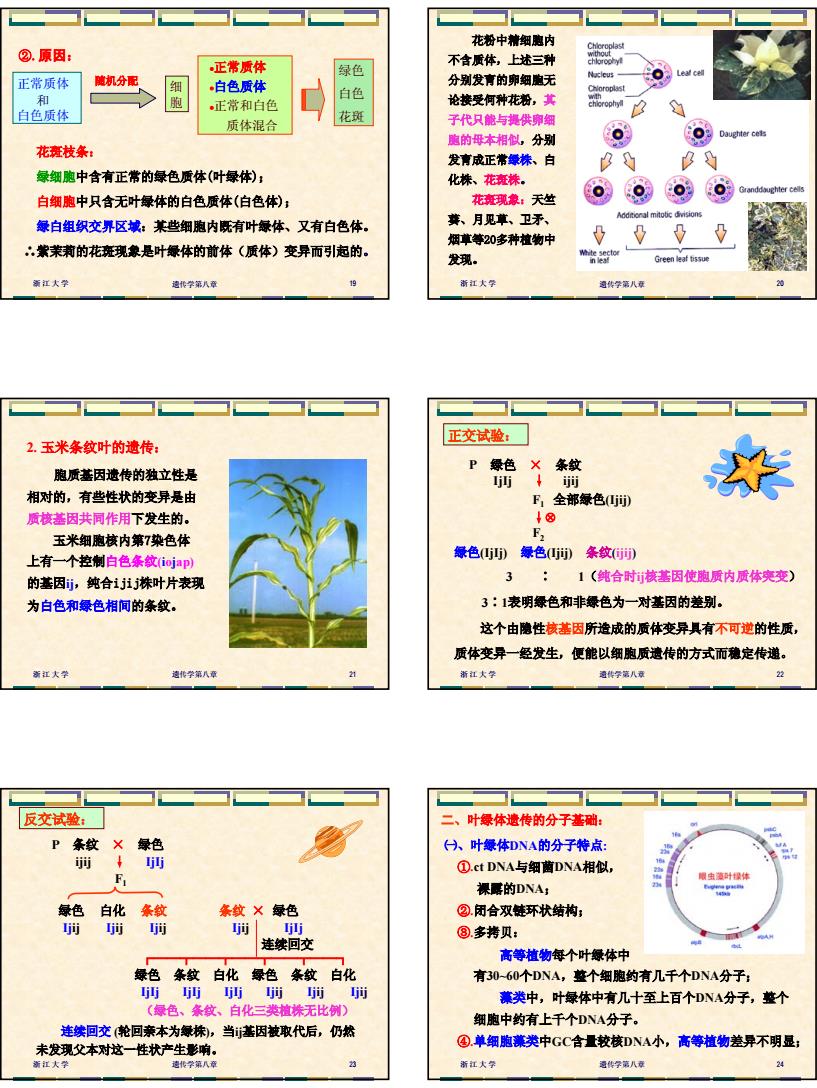
4 浙江大学 遗传学第八章 19 正常质体 和 白色质体 细 胞 •正常质体 •白色质体 •正常和白色 质体混合 随机分配 绿色 白色 花斑 花斑枝条: 绿细胞中含有正常的绿色质体(叶绿体); 白细胞中只含无叶绿体的白色质体(白色体); 绿白组织交界区域:某些细胞内既有叶绿体、又有白色体。 ∴紫茉莉的花斑现象是叶绿体的前体(质体)变异而引起的。 ②.原因: 浙江大学 遗传学第八章 20 花粉中精细胞内 不含质体,上述三种 分别发育的卵细胞无 论接受何种花粉,其 子代只能与提供卵细 胞的母本相似,分别 发育成正常绿株、白 化株、花斑株。 花斑现象:天竺 葵、月见草、卫矛、 烟草等20多种植物中 发现。 浙江大学 遗传学第八章 21 2. 玉米条纹叶的遗传: 胞质基因遗传的独立性是 相对的,有些性状的变异是由 质核基因共同作用下发生的。 玉米细胞核内第7染色体 上有一个控制白色条纹(iojap) 的基因ij,纯合ijij株叶片表现 为白色和绿色相间的条纹。 浙江大学 遗传学第八章 22 P 绿色 × 条纹 IjIj ↓ ijij F1 全部绿色(Ijij) ↓⊗ F2 绿色(IjIj) 绿色(Ijij) 条纹(ijij) 3 ∶ 1(纯合时ij核基因使胞质内质体突变) 3∶1表明绿色和非绿色为一对基因的差别。 这个由隐性核基因所造成的质体变异具有不可逆的性质, 质体变异一经发生,便能以细胞质遗传的方式而稳定传递。 正交试验: 浙江大学 遗传学第八章 23 P 条纹 × 绿色 ijij ↓ IjIj F1 绿色 白化 条纹 条纹 × 绿色 Ijij Ijij Ijij Ijij IjIj 连续回交 ┌──┬──┬──┬──┬──┐ 绿色 条纹 白化 绿色 条纹 白化 IjIj IjIj IjIj Ijij Ijij Ijij (绿色、条纹、白化三类植株无比例) 连续回交 (轮回亲本为绿株),当ij基因被取代后,仍然 未发现父本对这一性状产生影响。 反交试验: 浙江大学 遗传学第八章 24 ㈠、叶绿体DNA的分子特点: ①.ct DNA与细菌DNA相似, 裸露的DNA; ②.闭合双链环状结构; ③.多拷贝: 高等植物每个叶绿体中 有30~60个DNA,整个细胞约有几千个DNA分子; 藻类中,叶绿体中有几十至上百个DNA分子,整个 细胞中约有上千个DNA分子。 ④.单细胞藻类中GC含量较核DNA小,高等植物差异不明显; 二、叶绿体遗传的分子基础:
4 浙江大学 遗传学第八章 19 正常质体 和 白色质体 细 胞 •正常质体 •白色质体 •正常和白色 质体混合 随机分配 绿色 白色 花斑 花斑枝条: 绿细胞中含有正常的绿色质体(叶绿体); 白细胞中只含无叶绿体的白色质体(白色体); 绿白组织交界区域:某些细胞内既有叶绿体、又有白色体。 ∴紫茉莉的花斑现象是叶绿体的前体(质体)变异而引起的。 ②.原因: 浙江大学 遗传学第八章 20 花粉中精细胞内 不含质体,上述三种 分别发育的卵细胞无 论接受何种花粉,其 子代只能与提供卵细 胞的母本相似,分别 发育成正常绿株、白 化株、花斑株。 花斑现象:天竺 葵、月见草、卫矛、 烟草等20多种植物中 发现。 浙江大学 遗传学第八章 21 2. 玉米条纹叶的遗传: 胞质基因遗传的独立性是 相对的,有些性状的变异是由 质核基因共同作用下发生的。 玉米细胞核内第7染色体 上有一个控制白色条纹(iojap) 的基因ij,纯合ijij株叶片表现 为白色和绿色相间的条纹。 浙江大学 遗传学第八章 22 P 绿色 × 条纹 IjIj ↓ ijij F1 全部绿色(Ijij) ↓⊗ F2 绿色(IjIj) 绿色(Ijij) 条纹(ijij) 3 ∶ 1(纯合时ij核基因使胞质内质体突变) 3∶1表明绿色和非绿色为一对基因的差别。 这个由隐性核基因所造成的质体变异具有不可逆的性质, 质体变异一经发生,便能以细胞质遗传的方式而稳定传递。 正交试验: 浙江大学 遗传学第八章 23 P 条纹 × 绿色 ijij ↓ IjIj F1 绿色 白化 条纹 条纹 × 绿色 Ijij Ijij Ijij Ijij IjIj 连续回交 ┌──┬──┬──┬──┬──┐ 绿色 条纹 白化 绿色 条纹 白化 IjIj IjIj IjIj Ijij Ijij Ijij (绿色、条纹、白化三类植株无比例) 连续回交 (轮回亲本为绿株),当ij基因被取代后,仍然 未发现父本对这一性状产生影响。 反交试验: 浙江大学 遗传学第八章 24 ㈠、叶绿体DNA的分子特点: ①.ct DNA与细菌DNA相似, 裸露的DNA; ②.闭合双链环状结构; ③.多拷贝: 高等植物每个叶绿体中 有30~60个DNA,整个细胞约有几千个DNA分子; 藻类中,叶绿体中有几十至上百个DNA分子,整个 细胞中约有上千个DNA分子。 ④.单细胞藻类中GC含量较核DNA小,高等植物差异不明显; 二、叶绿体遗传的分子基础:
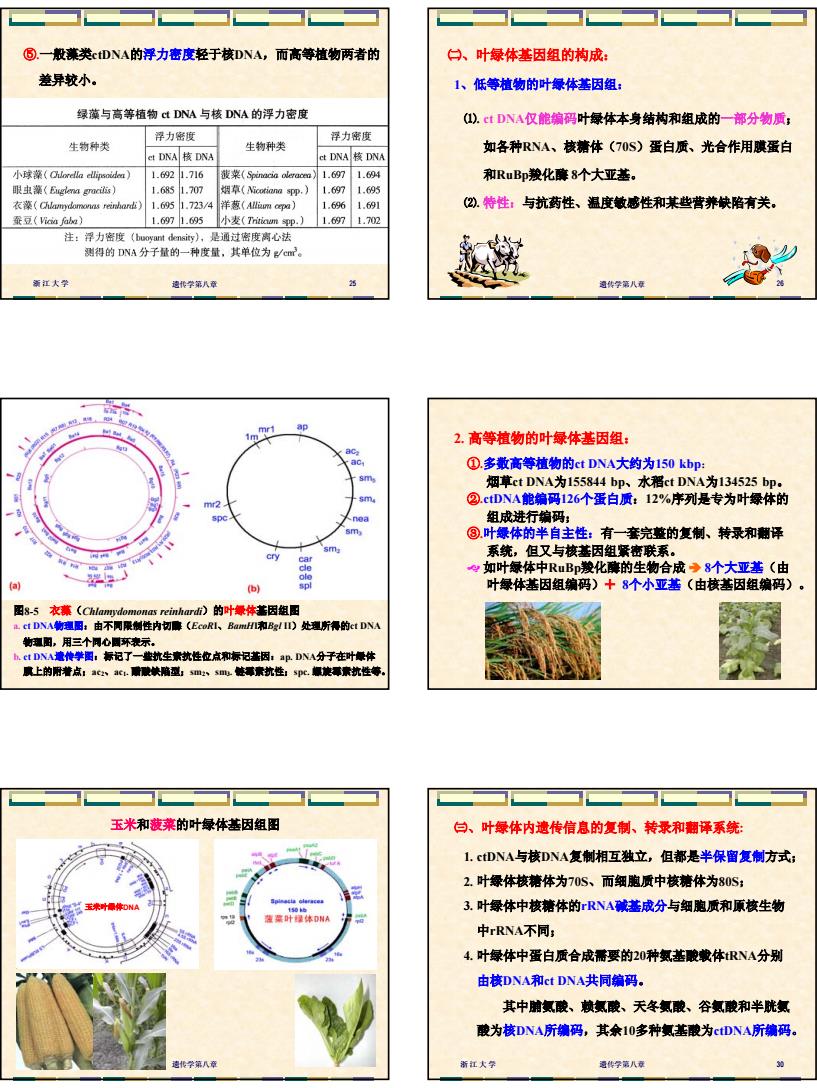
5 浙江大学 遗传学第八章 25 ⑤.一般藻类ctDNA的浮力密度轻于核DNA,而高等植物两者的 差异较小。 浙江大学 遗传学第八章 26 ㈡、叶绿体基因组的构成: 1、低等植物的叶绿体基因组: ⑴. ct DNA仅能编码叶绿体本身结构和组成的一部分物质; 如各种RNA、核糖体(70S)蛋白质、光合作用膜蛋白 和RuBp羧化酶 8个大亚基。 ⑵. 特性:与抗药性、温度敏感性和某些营养缺陷有关。 图8-5 衣藻(Chlamydomonas reinhardi)的叶绿体基因组图 a. ct DNA物理图:由不同限制性内切酶(EcoRI、BamHI和Bgl II)处理所得的ct DNA 物理图,用三个同心圆环表示。 b. ct DNA遗传学图:标记了一些抗生素抗性位点和标记基因:ap. DNA分子在叶绿体 膜上的附着点;ac2、ac1. 醋酸缺陷型;sm2、sm3. 链霉素抗性;spc. 螺旋霉素抗性等。 ①.多数高等植物的ct DNA大约为150 kbp: 烟草ct DNA为155844 bp、水稻ct DNA为134525 bp。 ②.ctDNA能编码126个蛋白质:12%序列是专为叶绿体的 组成进行编码; ③.叶绿体的半自主性:有一套完整的复制、转录和翻译 系统,但又与核基因组紧密联系。 ] 如叶绿体中RuBp羧化酶的生物合成 Î 8个大亚基(由 叶绿体基因组编码)ª 8个小亚基(由核基因组编码)。 2. 高等植物的叶绿体基因组: 浙江大学 遗传学第八章 29 玉米和菠菜的叶绿体基因组图 玉米叶绿体DNA 浙江大学 遗传学第八章 30 ㈢、叶绿体内遗传信息的复制、转录和翻译系统: 1. ctDNA与核DNA复制相互独立,但都是半保留复制方式; 2. 叶绿体核糖体为70S、而细胞质中核糖体为80S; 3. 叶绿体中核糖体的rRNA碱基成分与细胞质和原核生物 中rRNA不同; 4. 叶绿体中蛋白质合成需要的20种氨基酸载体tRNA分别 由核DNA和ct DNA共同编码。 其中脯氨酸、赖氨酸、天冬氨酸、谷氨酸和半胱氨 酸为核DNA所编码,其余10多种氨基酸为ctDNA所编码
5 浙江大学 遗传学第八章 25 ⑤.一般藻类ctDNA的浮力密度轻于核DNA,而高等植物两者的 差异较小。 浙江大学 遗传学第八章 26 ㈡、叶绿体基因组的构成: 1、低等植物的叶绿体基因组: ⑴. ct DNA仅能编码叶绿体本身结构和组成的一部分物质; 如各种RNA、核糖体(70S)蛋白质、光合作用膜蛋白 和RuBp羧化酶 8个大亚基。 ⑵. 特性:与抗药性、温度敏感性和某些营养缺陷有关。 图8-5 衣藻(Chlamydomonas reinhardi)的叶绿体基因组图 a. ct DNA物理图:由不同限制性内切酶(EcoRI、BamHI和Bgl II)处理所得的ct DNA 物理图,用三个同心圆环表示。 b. ct DNA遗传学图:标记了一些抗生素抗性位点和标记基因:ap. DNA分子在叶绿体 膜上的附着点;ac2、ac1. 醋酸缺陷型;sm2、sm3. 链霉素抗性;spc. 螺旋霉素抗性等。 ①.多数高等植物的ct DNA大约为150 kbp: 烟草ct DNA为155844 bp、水稻ct DNA为134525 bp。 ②.ctDNA能编码126个蛋白质:12%序列是专为叶绿体的 组成进行编码; ③.叶绿体的半自主性:有一套完整的复制、转录和翻译 系统,但又与核基因组紧密联系。 ] 如叶绿体中RuBp羧化酶的生物合成 Î 8个大亚基(由 叶绿体基因组编码)ª 8个小亚基(由核基因组编码)。 2. 高等植物的叶绿体基因组: 浙江大学 遗传学第八章 29 玉米和菠菜的叶绿体基因组图 玉米叶绿体DNA 浙江大学 遗传学第八章 30 ㈢、叶绿体内遗传信息的复制、转录和翻译系统: 1. ctDNA与核DNA复制相互独立,但都是半保留复制方式; 2. 叶绿体核糖体为70S、而细胞质中核糖体为80S; 3. 叶绿体中核糖体的rRNA碱基成分与细胞质和原核生物 中rRNA不同; 4. 叶绿体中蛋白质合成需要的20种氨基酸载体tRNA分别 由核DNA和ct DNA共同编码。 其中脯氨酸、赖氨酸、天冬氨酸、谷氨酸和半胱氨 酸为核DNA所编码,其余10多种氨基酸为ctDNA所编码