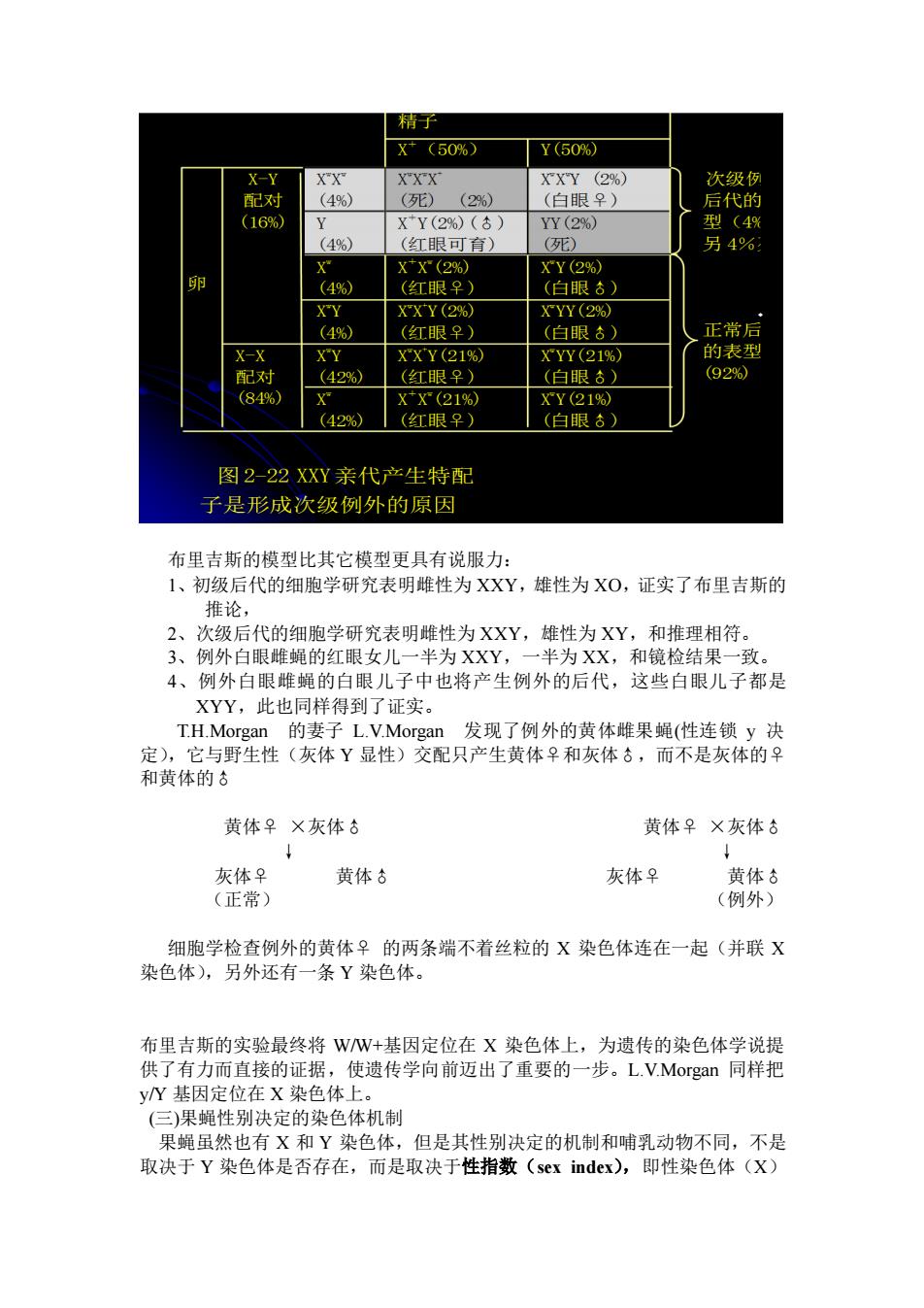
精于子 X(50%) Y(60% -Y XXX XXY (2%) 次级 配对 (4%) (29%) (白眼早) 后代的 (16% XY(2%)(s) YY(2%) 型(4% (4%) (工银可有) 死) 另4% XX(2% XY(2% %) (红眼早 (白眼) XY XX Y(2% XYY(2% (4%) (红眼早) (白眼】 正常后 -Y XXY(21% XYY(21% 配对 429%) (白眼】 (84% XX(21% XY(21% (42%) (红眼早) (白眼) 图2-22XXY亲代产生特配 子是形成次级例外的原因 布里吉斯的模型比其它模型更具有说服力: 1、初级后代的细胞学研究表明雌性为XXY 雄性为X0,证实了布里吉斯的 推论, 2、次级后代的细胞学研究表明雌性为XXY,雄性为XY,和推理相符。 3、例外白眼雌蝇的红眼女儿一半为XXY,一半为XX,和镜检结果一致。 4、例外白眼雌蝇的白眼儿子中也将产生例外的后代,这些白眼儿子都是 XYY,此也同样得到了证实。 T.H.Morgan的妻子L.V.Morgan 发现了例外的黄体雌果蝇(性连锁y决 定),它与野生性(灰体Y显性)交配只产生黄体♀和灰体♂,而不是灰体的♀ 和黄体的♂ 黄体♀×灰体 黄体♀×灰体 灰体♀ 黄体古 灰体♀ 黄体♂ (正常) (例外) 细胞学检查例外的黄体♀的两条端不着丝粒的X染色体连在一起(并联X 染色体),另外还有一条Y染色体 布里吉斯的实验最终将WW+基因定位在X染色体上,为遗传的染色体学说提 供了有力而直接的证据,使遗传学向前迈出了重要的一步。L.V.Morgan同样把 N基 定位在X染色体上 (三)果蝇性别决定的染色体机制 果蝇虽然也有X和Y染色体,但是其性别决定的机制和哺乳动物不同,不是 取决于Y染色体是否存在,而是取决于性指数(sex index),即性染色体(X)
布里吉斯的模型比其它模型更具有说服力: 1、初级后代的细胞学研究表明雌性为 XXY,雄性为 XO,证实了布里吉斯的 推论, 2、次级后代的细胞学研究表明雌性为 XXY,雄性为 XY,和推理相符。 3、例外白眼雌蝇的红眼女儿一半为 XXY,一半为 XX,和镜检结果一致。 4、例外白眼雌蝇的白眼儿子中也将产生例外的后代,这些白眼儿子都是 XYY,此也同样得到了证实。 T.H.Morgan 的妻子 L.V.Morgan 发现了例外的黄体雌果蝇(性连锁 y 决 定),它与野生性(灰体 Y 显性)交配只产生黄体♀和灰体♂,而不是灰体的♀ 和黄体的♂ 黄体♀ ×灰体♂ 黄体♀ ×灰体♂ ↓ ↓ 灰体♀ 黄体♂ 灰体♀ 黄体♂ (正常) (例外) 细胞学检查例外的黄体♀ 的两条端不着丝粒的 X 染色体连在一起(并联 X 染色体),另外还有一条 Y 染色体。 布里吉斯的实验最终将 W/W+基因定位在 X 染色体上,为遗传的染色体学说提 供了有力而直接的证据,使遗传学向前迈出了重要的一步。L.V.Morgan 同样把 y/Y 基因定位在 X 染色体上。 (三)果蝇性别决定的染色体机制 果蝇虽然也有 X 和 Y 染色体,但是其性别决定的机制和哺乳动物不同,不是 取决于 Y 染色体是否存在,而是取决于性指数(sex index),即性染色体(X)
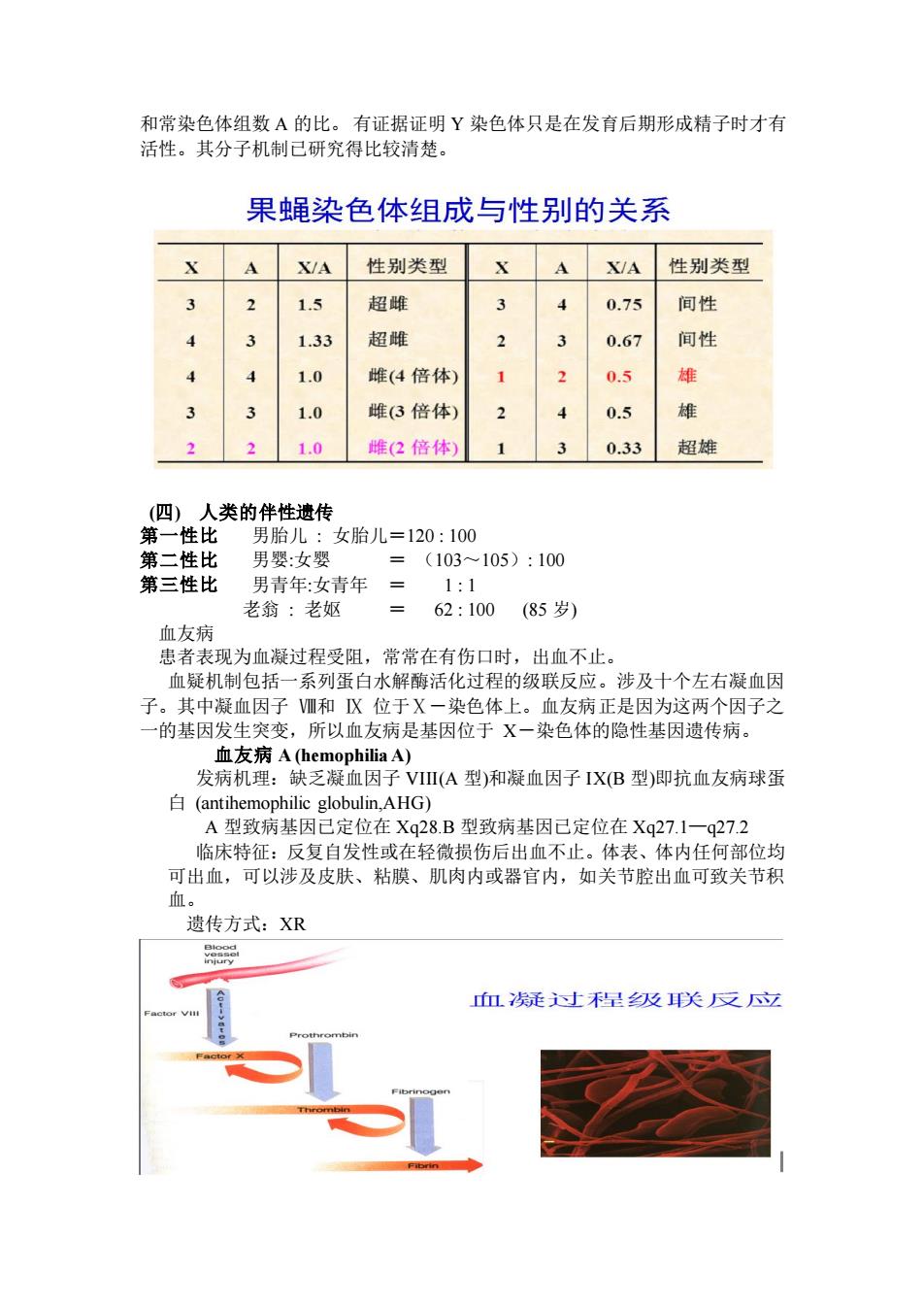
和常染色体组数A的比。有证据证明Y染色体只是在发有后期形成精子时才有 活性。其分子机制己研究得比较清楚。 果蝇染色体组成与性别的关系 A XyA性别类型 X AX/A 性别类型 1.5 超雌 3 4 0.75 间性 3 1.33 超雠 3 0.67 间性 1.0 雌(4倍体 2 0.5 雄 3 1.0 雌(3倍体) 0.5 雄 3 1.0 雌(2倍体 1 0.33 超雄 )人类的伴性遗传 第 一性比 男胎儿:女胎儿=120:100 第二性比 男:女 (103~105):100 第三性比 男青年:女青年= 1:1 老翁:老奴 =62:100(85岁) 血友病 患者表现为血凝过程受阻,常常在有伤口时,出血不止。 血疑机制包括一系列蛋白水解酶活化过程的级联反应。涉及十个左右凝血因 子。其中凝血因子Ⅷ和 X位于X一染色体上。血友病正是因为这两个因子之 一的基因发生突变,所以血友病是基因位于X一染色体的隐性基因遗传病。 血友病A(hemophilia A) 发病机理:缺乏凝血因子VII(A型)和凝血因子IX(B型)即抗血友病球蛋 白(antihem onhilic globulin AHg) A型致病基因己定位在Xq28B型致病基因已定位在Xq27.1一q272 临床特征:反复自发性或在轻微损伤后出血不止。体表、体内任何部位均 可出血,可以涉及皮肤、粘膜、肌肉内或器官内,如关节腔出血可致关节积 血。 遗传方式:XR 血凑凝过星及关反立
和常染色体组数 A 的比。 有证据证明 Y 染色体只是在发育后期形成精子时才有 活性。其分子机制已研究得比较清楚。 (四) 人类的伴性遗传 第一性比 男胎儿 女胎儿=120 100 第二性比 男婴女婴 = (103~105) 100 第三性比 男青年女青年 = 1 1 老翁 老妪 = 62 100 (85 岁) 血友病 患者表现为血凝过程受阻,常常在有伤口时,出血不止。 血疑机制包括一系列蛋白水解酶活化过程的级联反应。涉及十个左右凝血因 子。其中凝血因子 Ⅷ和 Ⅸ 位于Ⅹ-染色体上。血友病正是因为这两个因子之 一的基因发生突变,所以血友病是基因位于 X-染色体的隐性基因遗传病。 血友病 A (hemophilia A) 发病机理:缺乏凝血因子 VIII(A 型)和凝血因子 IX(B 型)即抗血友病球蛋 白 (antihemophilic globulin,AHG) A 型致病基因已定位在 Xq28.B 型致病基因已定位在 Xq27.1—q27.2 临床特征:反复自发性或在轻微损伤后出血不止。体表、体内任何部位均 可出血,可以涉及皮肤、粘膜、肌肉内或器官内,如关节腔出血可致关节积 血。 遗传方式:XR
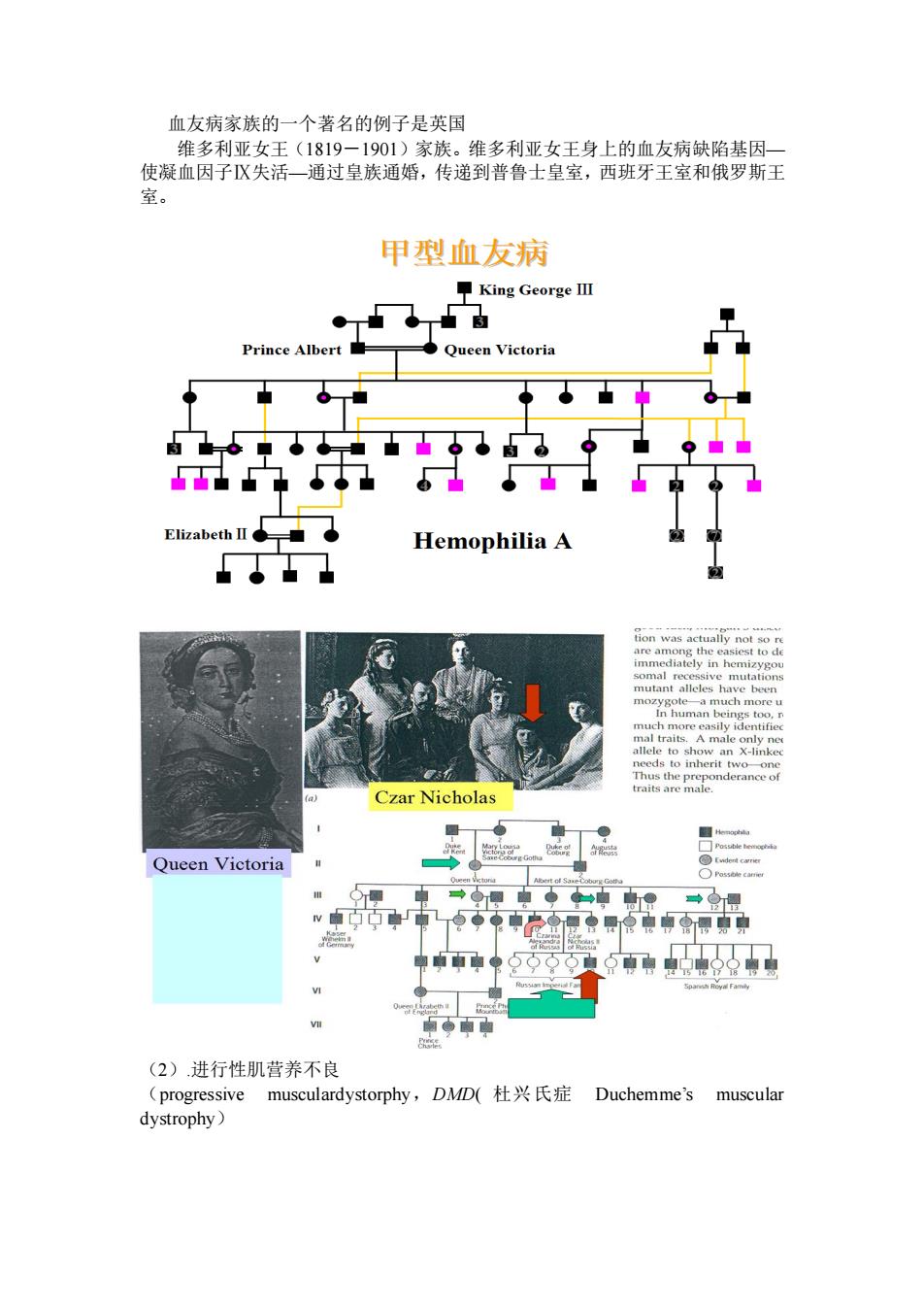
血友病家族的一个著名的例子是英国 维名利亚女王(1819一1901)家族。维多利亚女王身上的血友病缺陷基因 使凝血因子Ⅸ失活一通过皇族通婚,传递到普鲁士皇室,西班牙王室和俄罗斯王 甲型血友病 平King GeorgeⅢ Prince Albert Queen Victoria 中九p9 ElizabethⅡ◆ Hemophilia A Czar Nicholas Queen Victoria (2)讲行性肌营养不良 (progressive musculardystorphy,DMD(杜兴氏症Duchemme's muscular dystrophy
血友病家族的一个著名的例子是英国 维多利亚女王(1819-1901)家族。维多利亚女王身上的血友病缺陷基因— 使凝血因子Ⅸ失活—通过皇族通婚,传递到普鲁士皇室,西班牙王室和俄罗斯王 室。 (2).进行性肌营养不良 (progressive musculardystorphy,DMD( 杜兴氏症 Duchemme’s muscular dystrophy)

3.Y-连锁遗传 鱼类中背鳍上的斑点 外耳道多毛家族谣系 (三)其他伴性基因的遗传分析 (1)果蝇截毛基因(bb)的遗传 若雄性果蝇无Y染色体,则不育。原因:Y染色体上带有精子生成所必 需的生殖力基因 黑腹果蝇截毛基因(bb)的遗传: P早截刚毛×正常& ♀正常 截刚毛 XbX地 X'Y+ X'X' Xbyb F1♀正常×正常 早正常×正常 知Y 8±X XYb F2正常截刚毛正常正常 正常正常 正常截刚毛 X+X地XbX地X+Y*X地Y X'X+X+Xb X+Yb XbYb 果蝇截刚毛的“X和Y连锁”遗传方式 (2)芦花鸡的毛色遗传 ①.芦花基因B为显性,正常基因b为隐性,位于Z性染色体上。 ②W染色体上不带它的等位基因。 ③雄为 ZZ,雌鸡为ZW. 白、芦花鸡的毛色遗传: ①芦花基因B为显性,正常基因b为隐性,位于Z性染色体上。 ②W染色体上不带它的等位基因。 ③雄鸡为ZZ,难鸡为ZW
(三)其他伴性基因的遗传分析 (1)果蝇截毛基因(bb)的遗传 若雄性果蝇无 Y 染色体,则不育。原因:Y 染色体上带有精子生成所必 需的生殖力基因。 黑腹果蝇截毛基因(bb)的遗传: (2)芦花鸡的毛色遗传 ①. 芦花基因 B 为显性,正常基因 b 为隐性,位于 Z 性染色体上。 ② W 染色体上不带它的等位基因。 ③. 雄鸡为 ZZ,雌鸡为 ZW。 ㈢、芦花鸡的毛色遗传: ① 芦花基因 B 为显性,正常基因 b 为隐性,位于 Z 性染色体上。 ② W 染色体上不带它的等位基因。 ③ 雄鸡为 ZZ,雌鸡为 ZW

ZBW 芦花(雌) 正常(雄) 交叉遗传 ZbW 787b 正常(雌) 芦花(雄) 近亲繁殖 雌:雄 芦花:正常 ZeZ ZbZb 1”1 芦花(雄) 正常(雄) ZBW 芦花(雌) 正常(雌) × 正常(雌) 芦花(雄) ZBZb 芦花(雌) 芦花(雄) 近亲繁殖 鸡产花 ZBZb 芦花(雄) 芦花(雄) 芦花(雌) 正常(雌)