
第十一章数量遗传 教学内容要点: 第一节数量性状的特征及其遗传基础 第二节数量性状遗传研究的基本统计方法 第三节遗传率的估算及其应用 第四节近亲繁殖和杂种优势 前述的遗传现象是基于一个共同的遗传本质,即生物体的遗传表现直接由其基因型所决 定一可根据遗传群体的表现变异推测群体的基因型变异或基因的差异。质量性状(qualitative。 trai)的特点:表现型和基因型的变异不连续(discontinuous)。 在杂种后代的分离群体中→ ee1Y¥ 可采用经典遗传学分析方法, 研究其遗传动态。 e9/ 生物界中还存在另一类遗传性状,其表现型变异是连续的(continuous))e数量性状 (quantitative trait)。如,人身高、动物体重、生育期、果实大小,产量高低等。通过对表现 型变异的分析推断群体的遗传变异心借助数理统计的分析方法,可以有效地分析数量性状的 遗传规律 813 第一节数量性状的特征及其遗传基础
第十一章 数量遗传 教学内容要点: 第一节 数量性状的特征及其遗传基础 第二节 数量性状遗传研究的基本统计方法 第三节 遗传率的估算及其应用 第四节 近亲繁殖和杂种优势 前述的遗传现象是基于一个共同的遗传本质,即生物体的遗传表现直接由其基因型所决 定→可根据遗传群体的表现变异推测群体的基因型变异或基因的差异。质量性状(qualitative trait)的特点:表现型和基因型的变异不连续(discontinuous)。 生物界中还存在另一类遗传性状,其表现型变异是连续的(continuous)è数量性状 (quantitative trait)。如,人身高、动物体重、生育期、果实大小,产量高低等。通过对表现 型变异的分析推断群体的遗传变异 è借助数理统计的分析方法,可以有效地分析数量性状的 遗传规律。 第一节 数量性状的特征及其遗传基础
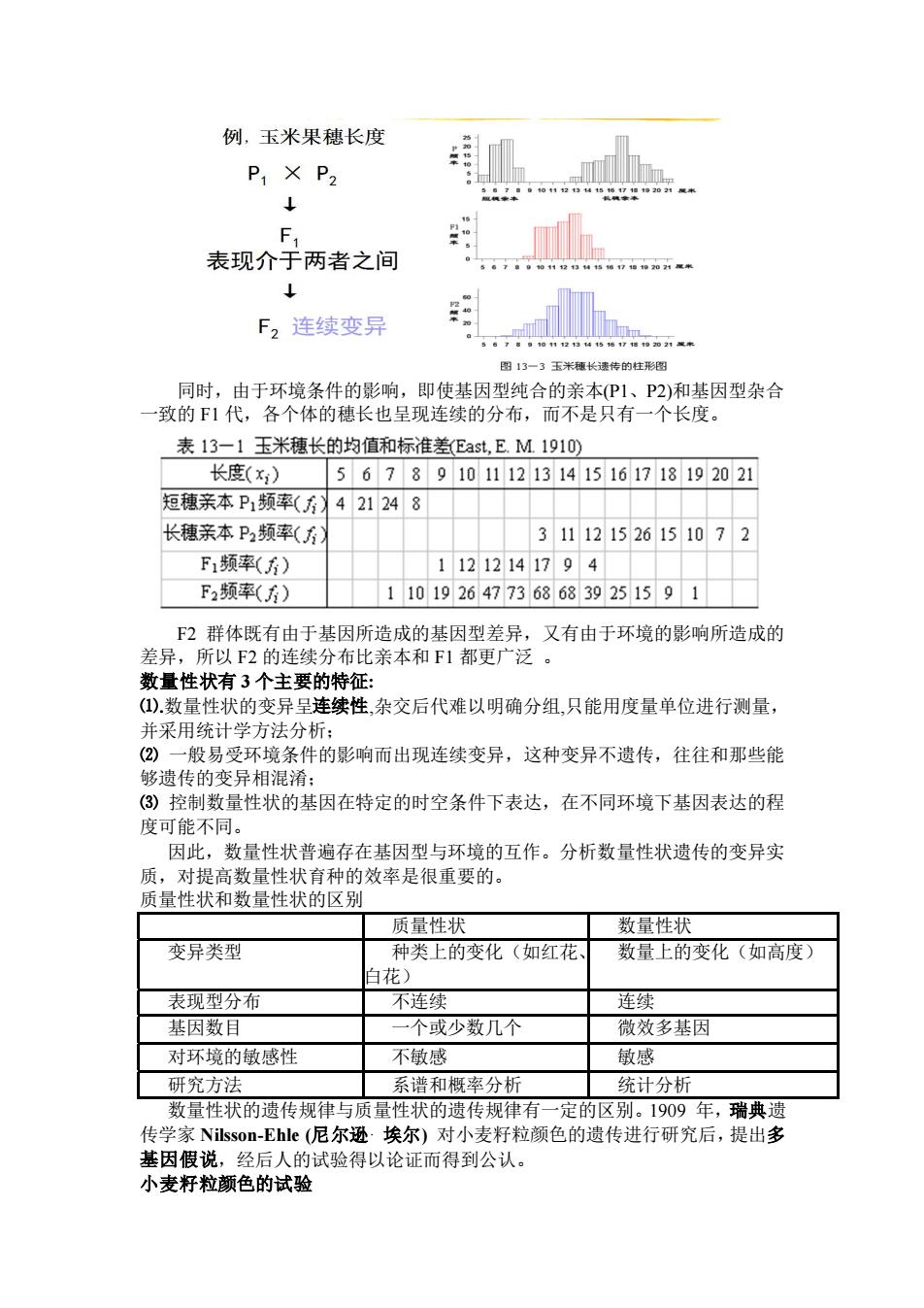
例,玉米果穗长度 P1×P2 F. 表现介于两者之间 F2连续变异 图13一3玉米嫩长速传的柱形图 同时,由于环境条件的影响,即使基因型纯合的亲本(P1、P2)和基因型杂合 一致的F1代,各个体的穗长也呈现连续的分布,而不是只有一个长度。 表13一1玉米穗长的均值和标准差Eat,EM1910) 长度(x) 56789101112131415161718192021 短穗亲本P1频率(方421248 长穗亲本P2频率(方) 311121526151072 F1频率(6) 11212141794 F2频率() 11019264773686839251591 F2群体既有由于基因所造成的基因型差异,又有由于环境的影响所造成的 差异,所以F2的连续分布比亲本和F1都更广泛。 新最性状有3个士要的特征 ().数量性 的变异呈连续性,杂交后代难以明确分组,只能用度量单位进行测量, 并采用统计学方法分析: (2)一般易受环境条件的影响而出现连续变异,这种变异不遗传,往往和那些能 够遗传的变异相混淆: (3)控制数量性状的基因在特定的时空条件下表达,在不同环境下基因表达的程 度可能不同。 因此,数量性状普遍存在基因型与环境的互作。分析数量性状遗传的变异实 质,对提高数量性状育种的效率是很重要的。 质量性状和数量性状的区别 质量性状 数量性状 变异类型 种类上的变化(如红花 数匣上的变化(如高度) 白花) 表现型分宿 不库续 车续 基因数目 一个或少数几个 微效多基因 对环境的敏成性 不敏感 鼓感 研究方法 系谱和概率分新 统计分析 数量性状的遗传规律与质量性状的遗传规律有一定的区别。1909年,瑞典遗 传学家Nilsson-Ehle(尼尔逊埃尔)对小麦籽粒颜色的遗传进行研究后,提出多 基因假说,经后人 试验得以论证而得到公认。 小麦籽粒颜色的试验
同时,由于环境条件的影响,即使基因型纯合的亲本(P1、P2)和基因型杂合 一致的 F1 代,各个体的穗长也呈现连续的分布,而不是只有一个长度。 F2 群体既有由于基因所造成的基因型差异,又有由于环境的影响所造成的 差异,所以 F2 的连续分布比亲本和 F1 都更广泛 。 数量性状有 3 个主要的特征: ⑴.数量性状的变异呈连续性,杂交后代难以明确分组,只能用度量单位进行测量, 并采用统计学方法分析; ⑵ 一般易受环境条件的影响而出现连续变异,这种变异不遗传,往往和那些能 够遗传的变异相混淆; ⑶ 控制数量性状的基因在特定的时空条件下表达,在不同环境下基因表达的程 度可能不同。 因此,数量性状普遍存在基因型与环境的互作。分析数量性状遗传的变异实 质,对提高数量性状育种的效率是很重要的。 质量性状和数量性状的区别 质量性状 数量性状 变异类型 种类上的变化(如红花、 白花) 数量上的变化(如高度) 表现型分布 不连续 连续 基因数目 一个或少数几个 微效多基因 对环境的敏感性 不敏感 敏感 研究方法 系谱和概率分析 统计分析 数量性状的遗传规律与质量性状的遗传规律有一定的区别。1909 年,瑞典遗 传学家 Nilsson-Ehle (尼尔逊· 埃尔) 对小麦籽粒颜色的遗传进行研究后,提出多 基因假说,经后人的试验得以论证而得到公认。 小麦籽粒颜色的试验

红粒×白粒 F1为中间型,不能区别显、隐性: F2则由红色到白色, 表现各种不同程度的类型。 按照他的解释,数量性状是许多彼此独立的基因作用的结果,各基因对性状 表现的效果较微,但其遗传方式仍然服从孟德尔遗传规律。 多基因假说要点 1.决定数量性状的基因数目很多: 2.各基因的效应相等; 3.各个等位基因的表现为不完全显性或无显性或有增效和减效作用: 4.各基因的作用是累加性的。 数量性状的深入研究已进一步丰富和发展了多基因假说。最简单的数量性 状,可假定由2对或3对基因决定的。现将小麦籽粒颜色2对和3对基因的遗传 动态图示如下: (①)小麦籽粒颜色受2对重叠基因决定时的遗传动态 P R 白粒 rinran 个 红粒 RifRara F2 ® 表现型类别 红色 表现型比例 深红中深红中红淡红一白色 4 红粒有放基因数 4R 3R IR OR 基因型 R ■红粒:白粒 11 (2)小麦籽粒颜色受三对重叠基因决定时的遗传动态
按照他的解释,数量性状是许多彼此独立的基因作用的结果,各基因对性状 表现的效果较微,但其遗传方式仍然服从孟德尔遗传规律。 多基因假说要点 1. 决定数量性状的基因数目很多; 2. 各基因的效应相等; 3. 各个等位基因的表现为不完全显性或无显性 或有增效和减效作用; 4. 各基因的作用是累加性的。 数量性状的深入研究已进一步丰富和发展了多基因假说。最简单的数量性 状,可假定由 2 对或 3 对基因决定的。现将小麦籽粒颜色 2 对和 3 对基因的遗传 动态图示如下: (1)小麦籽粒颜色受 2 对重叠基因决定时的遗传动态 (2)小麦籽粒颜色受三对重叠基因决定时的遗传动态

白粒 Y1YY2T2 Iyry F1 红粒 RinRata Rary F2 1⊙ 表现型类别 红色 最深红暗红深红 中深红 中红淡红白色 表现型比例 615 20 1561 红粒有效基因数 6R SR 4R. 3R 2R 1R OR 红粒:白粒 63:1 当性状由n对独立基因决定时,则F2的表现型频率为: (经R+2)2 在自然界,数量性状的遗传方式要复杂得多,往往由多对基因决定:且经常 受到环境条件的作用,使遗传的和不遗传的变异混在一起,不易区别开来。所以, 对于数量性状的研究 一定要采用统计学的分析方法。近年来,借助数量性状基 因位点(quantitative trait oc,QTL)和分子标记作图技术,已经可以在分子 标记连锁图上标出单个基因位点的位置、并确定其基因效应。 主基因(major gene)和微效基因(minor gene)数量性状可以由少数效应较大的 主基因控制,也可由数目较多、效应较小的微效多基因或微效基因所控制。 主基因:控制某个性状表现的效应较大的少数基因:微效基因:数目较多 但每个基因对表现型的影响较小:修饰基因(modifying gene): 一些性状虽主 要由少数主基因控制,但另外还存在 一些效应微小的修饰基因,其作用微小,但 能增强或削弱主基因对基因型的作用。 微效基因的效应类型 各个微效基因的遗传效应值不尽相等,效应的类型包括等位基因的加性效应 additive effect,A)、显性效应(d inance effect,D),以及非等位基因间的上位 性效应(epitasiseffect,.I),还包括这些基因主效应与环境的互作效应。 P=G+E+GE ·P=A+D+I+E+GE 超亲遗传(r nsgressive inheritan e) 在植物杂交时,杂种后代出现的一种超越双亲现象 如水稻的两个品种: 早熟(A2A2B2B2C1C1)×晚熟(A1A1B1B1C2C2) FI (A1A2B1B2C1C2)熟期介于双亲之间 27种基因型 (其中A1A1B1B1C1C1的个体将比晚熟亲本更晚 而A2A2B2B2C2C2的个体将比早熟亲本更早)
当性状由 n 对独立基因决定时,则 F2 的表现型频率为: 在自然界,数量性状的遗传方式要复杂得多,往往由多对基因决定;且经常 受到环境条件的作用,使遗传的和不遗传的变异混在一起,不易区别开来。所以, 对于数量性状的研究,一定要采用统计学的分析方法。近年来,借助数量性状基 因位点(quantitative trait loci,QTL)和分子标记作图技术,已经可以在分子 标记连锁图上标出单个基因位点的位置、并确定其基因效应。 主基因(major gene)和微效基因(minor gene):数量性状可以由少数效应较大的 主基因控制,也可由数目较多、效应较小的微效多基因或微效基因所控制。 主基因:控制某个性状表现的效应较大的少数基因;微效基因:数目较多, 但每个基因对表现型的影响较小;修饰基因(modifying gene): 有一些性状虽主 要由少数主基因控制,但另外还存在一些效应微小的修饰基因,其作用微小,但 能增强或削弱主基因对基因型的作用。 微效基因的效应类型 各个微效基因的遗传效应值不尽相等,效应的类型包括等位基因的加性效应 (additive effect,A)、显性效应(dominance effect,D) ,以及非等位基因间的上位 性效应(epitasis effect,I ),还包括这些基因主效应与环境的互作效应。 超亲遗传(transgressive inheritance) : 在植物杂交时,杂种后代出现的一种超越双亲现象。 如 水稻的两个品种: P 早熟(A2A2B2B2C1C1) × 晚熟(A1A1B1B1C2C2) ↓ F1 (A1A2B1B2C1C2) 熟期介于双亲之间 ↓ F2 27 种基因型 (其中 A1A1B1B1C1C1 的个体将比晚熟亲本更晚, 而 A2A2B2B2C2C2 的个体将比早熟亲本更早)

第二节数量性状遗传研究的基本统计方法 数量遗传学一研究数量性状在群体内的遗传规律一目的:将总表现型变异分 解为遗传和非遗传部分。统计参数:均值(Mean)、方差(Variance)、协方之 (Covariance)和相关系数等→提供可以解释遗传变异和预测变异在下一代表现程 度所需的信息。 “在研究数量性状的遗传规律时,需用数理统计的方法。 第三节遗传率的估算及其应用 一、遗传率的概念 二、遗传率的估算 三、遗传率在育种上的应用 一、遗传率的概念 L.遗传率或遗传力山eritability:)指遗传方差(VG)在总方差(P)中所占比值,可 作为杂种后代进行选择的一个指标。是度量性状的遗传变异占表现型变异相对比 率的重要遗传参数。 广义遗传率heritability in the broad sense,H2) 狭义遗传率heritability in the narrow sense,h2) 简单的数最贵传分析, 般假定遗传效应只包括加性效应和显性效应,且不存在 基因效应×环境效应的互作,表现型方差简单地分解为: VP= VG+Ve VA+VD+Ve 广义遗传率(heritability in the broad sense,H2)定义为总的遗传方差占表现型方 差的比率 H2=VG/VP =(VA+VD)/(VA+VD+Ve) 狭义遗传率(heritability in the narrow sense,h2)定义为加性遗传方差占表现型方 差的比率。 h2=VA/VP VA/(VA+VD+Ve) 2.导致群体表现型发生变异的遗传原因 ①遗传主效应产生的普通遗传变异,由遗传方差(G)来度量: ②.基因型×环境的互作效应产生的互作遗传变异, 由基因型×环境的互作方差(VGE)来度量。 遗传率也可分解为二个分量: 普通遗传率(general heritability) 互作遗传率(interaction heritability)。 这种分解适用于广义遗传率(H2)和狭义遗传率(2)。 3.遗传率的组成 ①广义遗传率(H2)的组成: H2=HG2+HGE2 其中: HG2是普通广义遗传率(general heritability in broad sense),定义为遗传方差占表 现型方差的比率,HG2=VGVP:
第二节 数量性状遗传研究的基本统计方法 数量遗传学→研究数量性状在群体内的遗传规律→目的:将总表现型变异分 解为遗传和非遗传部分。统计参数:均值(Mean)、方差(Variance)、协方差 (Covariance)和相关系数等→提供可以解释遗传变异和预测变异在下一代表现程 度所需的信息。 ∴在研究数量性状的遗传规律时,需用数理统计的方法。 第三节 遗传率的估算及其应用 一、遗传率的概念 二、遗传率的估算 三、遗传率在育种上的应用 一、遗传率的概念 1. 遗传率或遗传力(heritability):指遗传方差(VG)在总方差(VP)中所占比值,可 作为杂种后代进行选择的一个指标。是度量性状的遗传变异占表现型变异相对比 率的重要遗传参数。 广义遗传率(heritability in the broad sense,H2) 狭义遗传率(heritability in the narrow sense,h2) 简单的数量遗传分析,一般假定遗传效应只包括加性效应和显性效应,且不存在 基因效应×环境效应的互作,表现型方差简单地分解为: VP = VG + Ve = VA + VD + Ve 广义遗传率(heritability in the broad sense,H2)定义为总的遗传方差占表现型方 差的比率. H 2 =VG/VP =(VA+VD)/(VA+VD+Ve) 狭义遗传率(heritability in the narrow sense,h2)定义为加性遗传方差占表现型方 差的比率。 h2 = VA/VP =VA/(VA+VD+Ve) 2.导致群体表现型发生变异的遗传原因 ①.遗传主效应产生的普通遗传变异,由遗传方差(VG)来度量; ②.基因型×环境的互作效应产生的互作遗传变异, 由基因型×环境的互作方差(VGE)来度量。 遗传率也可分解为二个分量: 普通遗传率(general heritability) 互作遗传率(interaction heritability)。 这种分解适用于广义遗传率(H2)和狭义遗传率(h2)。 3.遗传率的组成 ①.广义遗传率(H2)的组成: H2 = HG2 + HGE2 其中: HG2 是普通广义遗传率(general heritability in broad sense), 定义为遗传方差占表 现型方差的比率,HG2=VG/VP;