
第七章基因精细结构遗传分析 教学内容 第一节基因概念 第二节重组测验 第三节互补测验 第四节缺失作图 第五节断裂基因与重叠基因 第六节基因功能 教学内容要点 基因的概念、重组测验、互补测验、 断裂基因与重叠基因、基因的功能 难点 重组测验、互补测验与基因的功能 7.1基因概念和精细结构 7.1.1基因概念的发展 (1)遗传因子(孟德尔) (2)染色体是基因载体(摩尔根〉 (3)DNA是遗传物质(肺炎双球菌的转化实验) (4)基因是有功能的DNA片段(G.Beadle和E.Tatum:一基因一酶) (⑤)操纵子模型(Jacob和Monod) (6)跳跃基因和断裂基因的发现 表了-1限成中特移植:哈 供体受体移描用的颈色 1 鲜红 鲜红 (33 cn 群红 4) 鲜红 5ù 6 鲜红 7.1.2基因的类别和相互关系 (1)结构基因 (2)rRNA和tRNA基因 (3)启动子和操纵基因 7.1.2基因的类别及其相互关系
第七章 基因精细结构遗传分析 教学内容 第一节 基因概念 第二节 重组测验 第三节 互补测验 第四节 缺失作图 第五节 断裂基因与重叠基因 第六节 基因功能 教学内容要点 基因的概念、重组测验、互补测验、 断裂基因与重叠基因、基因的功能 难点 重组测验、互补测验与基因的功能 7.1 基因概念和精细结构 7.1.1 基因概念的发展 (1)遗传因子(孟德尔) (2)染色体是基因载体(摩尔根) (3)DNA 是遗传物质(肺炎双球菌的转化实验) (4)基因是有功能的 DNA 片段(G.Beadle 和 E. Tatum:一基因一酶) (5)操纵子模型(Jacob 和 Monod) (6)跳跃基因和断裂基因的发现 7.1.2 基因的类别和相互关系 (1)结构基因 (2)rRNA 和 tRNA 基因 (3)启动子和操纵基因 7.1.2 基因的类别及其相互关系 表 7-1 眼成虫盘移植实验 供体 受体 移植眼的颜色 (1) + ν 鲜红 (2) ν + 鲜红 (3) + cn 鲜红 (4) cn + 鲜红 (5) cn ν 橙红 (6) ν cn 鲜红 + 鲜红的(bright red),v:桔黄色眼(vermilion), c n 橙红色眼(cimnabar),和暗红色眼(scarlet)

根据基因的功能和性质,分为: (一)结构基因(structural genes)与调节基因(regulatory genes) (二)核糖体RM基因(ribosomal RNA genes,简称rDNA)与转移RNA (transfer RNAgenes,.简称tDNA) (三)启动子(promotor)与操纵基因(operator) 前者是转录时RNA聚合酶与DNA结合的部位:后者是调节基因产物阻遏蛋 白或激活蛋白与DNA结合的部位。 7.1.3基因与DN 多数肽链由150~300个氨基酸组成,按三联密码子,须有450~ 900个核苷酸对编码,加上基因内不编码的核苷酸序列,一个基因约 有500~6000个核苷酸对。并非DNA分子上任一含有几千个核苷酸 对的区段都是一个基因,基因是一个含有特定遗传信息的DNA分子区 段。特定核苷酸序列与其转录产物RNA核苷酸序列或翻译产物多肽 链氨基酸序列相对应就是基因,要同时测定某一段DNA的核苷酸序列 和相应产物的序列。 7.2 重组测验 7.2.1拟等位基因 黑腹果蝇红眼由一个显性基因控制,位于X染色体上,果蝇眼睛还 有许多其他颜色,如粉红色、杏色、伊红、象牙色和白色等突变型。 早期研究认为控制这些眼晴颜色性状的基因是等位的,之间是复等 位关系。用“十”代表野生型红色眼基因,W代表杏色眼基因,W代 表白色眼基因,当杏色眼(Wa/wa)与白眼(W/Y)果蝇杂交时, F1为杏色眼,如果w阳与w是等位基因,F1应只有两种亲本表型,但 在大量F1群体中约有1/1000野生型红眼果蝇。红眼的出现不是突 变,因为突变没有如此高频率。 进一步研究证明杏色眼基因和白眼基因虽然在染色体上所占位置相 同,即位于同一基因,但属于不同位点,之间可发生交换。 如杏色眼wa++/wa+x白眼w/Y,F1杏色眼。wa+/+W和wa+/Y, F1雌蝇减数分裂时发生交换形成+与Ww配子,+配子与任何其他配 子结合所形成的F1个体均表现为野生型红眼果蝇,因而F1群体中出 现野牛型个体。 对杏色限Wa+/+W与野生型++/W阳w两种个体进行比较发现,基因 组成一样,但排列不同,前者两个突变分别在两条染色体上,为反式
根据基因的功能和性质,分为: (一)结构基因(structural genes)与调节基因(regulatory genes) (二)核糖体 RNA 基因(ribosomal RNA genes,简称 rDNA)与转移 RNA (transfer RNAgenes,简称 tDNA) (三)启动子(promotor)与操纵基因(operator) 前者是转录时 RNA 聚合酶与 DNA 结合的部位;后者是调节基因产物阻遏蛋 白或激活蛋白与 DNA 结合的部位。 7.1.3 基因与 DNA 多数肽链由 150~ 300 个氨基酸组成,按三联密码子,须有 450~ 900 个核苷酸对编码,加上基因内不编码的核苷酸序列,一个基因约 有 500~6 000 个核苷酸对。并非 DNA 分子上任一含有几千个核苷酸 对的区段都是一个基因,基因是一个含有特定遗传信息的 DNA 分子区 段。 特定核苷酸序列与其转录产物 RNA 核苷酸序列或翻译产物多肽 链氨基酸序列相对应就是基因,要同时测定某一段 DNA 的核苷酸序列 和相应产物的序列。 7.2 重组测验 7.2.1 拟等位基因 黑腹果蝇红眼由一个显性基因控制,位于 X 染色体上,果蝇眼睛还 有许多其他颜色,如粉红色、杏色、伊红、象牙色和白色等突变型。 早期研究认为控制这些眼睛颜色性状的基因是等位的,之间是复等 位关系。用“+”代表野生型红色眼基因,wa 代表杏色眼基因,w 代 表白色眼基因,当杏色眼( w a/ wa )与白眼(w /Y)果蝇杂交时, F1 为杏色眼,如果 wa 与 w 是等位基因,F1应只有两种亲本表型,但 在大量 F1 群体中约有 1/1000 野生型红眼果蝇。红眼的出现不是突 变,因为突变没有如此高频率。 进一步研究证明杏色眼基因和白眼基因虽然在染色体上所占位置相 同,即位于同一基因,但属于不同位点,之间可发生交换。 如杏色眼 wa+ + /wa + x 白眼 w/Y,F1 杏色眼。wa +/+ w 和 w a +/Y, F1 雌蝇减数分裂时发生交换形成++与 waw 配子,++配子与任何其他配 子结合所形成的 F1 个体均表现为野生型红眼果蝇,因而 F1 群体中出 现野生型个体。 对杏色眼 wa+/+w 与野生型++/ wa w 两种个体进行比较发现,基因 组成一样,但排列不同,前者两个突变分别在两条染色体上,为反式

(trans)排列,后者则是两个突变同时排在一条染色体上,而另一条 染色体上两个位点均正常,为顺式(cis)排列。 反式排列表现为突变型,顺式排列为野生型。这种由于排列方式不 同而表型不同的现象称为顺反位置效应(cis-trans position effects),并将这种紧密连锁的功能性等位基因,但不是结构性的等 位基因称为拟等位基因(pseudoallele)。拟等位基因的发现证明了 基因的可分性。 7.2.2噬菌体突变型 S.Benzer对大肠杆菌噬菌体T4的rIIA和rIIB两个基因的结构 分析,证明了基因的可分性,基因内有大量的突变子和重组子。 噬菌体的突变型可归为: ①噬菌斑形态突变型:一些是由于侵染寄主后溶菌速度的快慢而形成 大小不同的噬菌斑(plaque):另一些是由于被感染细菌是全部或是 部分被杀死而形成清晰或混浊的噬菌斑。 ②寄主范围突变型: 噬菌体感染细菌时,首先吸附于细胞表面专一受体上,由受体基因控 制,如果受体发生改变,可能使噬菌体不能附着,从而该噬菌体的寄 主范围缩小。另外噬菌体突变也可扩大寄生范围。因为决定噬菌斑形 态和宿主范围突变的基因在其基因组中相当狭窄的特定区段里,大多 数基因涉及生命过程必不可少的功能,所以上述突变通常是致死的。 ③条件致死突变型: 条件致死突变型在遗传学研究中具重要意义,通过这种突变已鉴定出噬 菌体的大部分基因 Benzer所用T4的rII突变就是责传学研究中所用的第个条件改 死突变型。 T4噬菌体有多个迅速裂解突变型,分别称为rl,rIL,rIII等 它们位于染色体DNA的不同区段,这3组突变型由于在大肠杆菌不同 菌株上的反应不同可以相互区别 T4rIⅡ突变使所侵染细胞迅速裂解形成大噬菌斑,所以称为rII 突变型 Benzer对rII区域的突变进行了分析:rII突变感染大肠杆菌B菌株 后迅速裂解,形成比野生型大的噬菌斑,从而从大量的r11+中筛选出 r1。另外r11突变型感染带有原噬菌体的大肠杆菌K(入)菌株时, 不产生子代,而野生型 T4rII在大肠杆菌K(入)菌株中能正常增
(trans)排列,后者则是两个突变同时排在一条染色体上,而另一条 染色体上两个位点均正常,为顺式(cis)排列。 反式排列表现为突变型,顺式排列为野生型。这种由于排列方式不 同 而 表型 不同 的现 象 称为 顺反 位置 效 应( cis-trans position effects),并将这种紧密连锁的功能性等位基因,但不是结构性的等 位基因称为拟等位基因(pseudoallele)。拟等位基因的发现证明了 基因的可分性。 7.2.2 噬菌体突变型 S.Benzer 对大肠杆菌噬菌体 T4 的 r IIA 和 r IIB 两个基因的结构 分析,证明了基因的可分性,基因内有大量的突变子和重组子。 噬菌体的突变型可归为: ①噬菌斑形态突变型:一些是由于侵染寄主后溶菌速度的快慢而形成 大小不同的噬菌斑(plaque);另一些是由于被感染细菌是全部或是 部分被杀死而形成清晰或混浊的噬菌斑。 ②寄主范围突变型: 噬菌体感染细菌时,首先吸附于细胞表面专一受体上,由受体基因控 制,如果受体发生改变,可能使噬菌体不能附着,从而该噬菌体的寄 主范围缩小。另外噬菌体突变也可扩大寄生范围。因为决定噬菌斑形 态和宿主范围突变的基因在其基因组中相当狭窄的特定区段里,大多 数基因涉及生命过程必不可少的功能,所以上述突变通常是致死的。 ③条件致死突变型: 条件致死突变型在遗传学研究中具重要意义,通过这种突变已鉴定出噬 菌体的大部分基因。 Benzer 所用 T4 的 r II 突变就是遗传学研究中所用的第一个条件致 死突变型。 T4 噬菌体有多个迅速裂解突变型,分别称为 rl, r II,rIII 等, 它们位于染色体 DNA 的不同区段,这 3 组突变型由于在大肠杆菌不同 菌株上的反应不同可以相互区别。 T4 r II 突变使所侵染细胞迅速裂解形成大噬菌斑,所以称为 r II 突变型。 Benzer 对 r II 区域的突变进行了分析: r II 突变感染大肠杆菌 B 菌株 后迅速裂解,形成比野生型大的噬菌斑,从而从大量的 rll+中筛选出 rll。另外 rll 突变型感染带有原噬菌体的大肠杆菌 K(λ)菌株时, 不产生子代,而野生型 T4 rII 在大肠杆菌 K(λ)菌株中能正常增
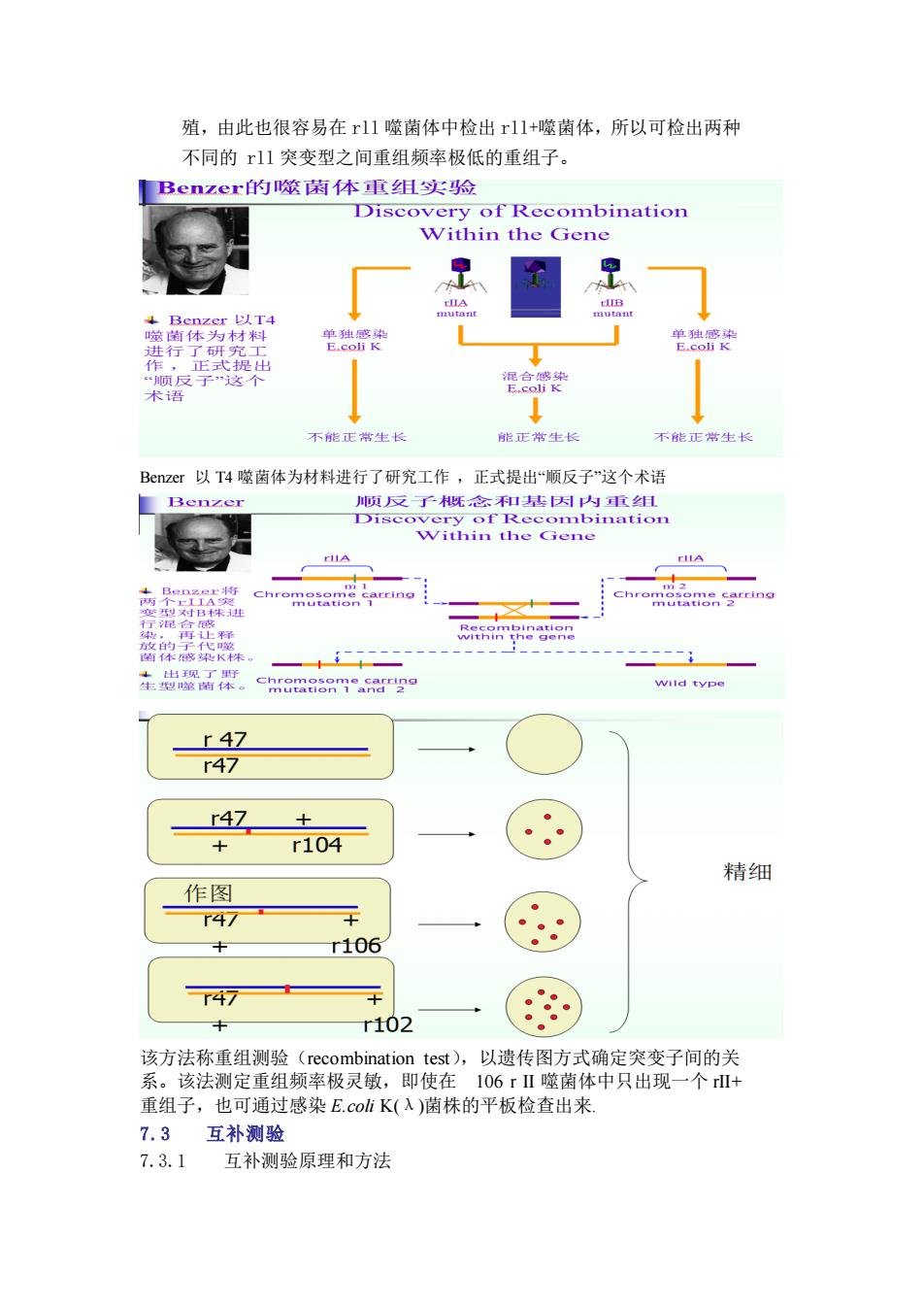
殖,由此也很容易在r11噬菌体中检出r11+噬菌体,所以可检出两种 不同的r1突变型之间重组频率极低的重组子。 Benzer的噬菊体重组实险 Discovery of Recombination Within the Gene 克B 以4 行 单独 不能正生 Benzer以T4噬菌体为材料进行了研究工作,正式提出“顺反子"这个术语 Benzer 顺反子既念和基因内重组 生出 a8o3.29 typ r4☑ r47 r47 r104 精细 作图 47 r106 r47 r102 该方法称重组测哈(rec mbination est),以遗传图方式确定突变子间的关 系 该法测定重组频率极灵敏,即使在 106rⅡ噬菌体中只出现 个 + 重组子,也可通过感染E.cohK(入)菌株的平板检查出来 7.3互补测验 7.3.1 互补测验原理和方法
殖,由此也很容易在 rll 噬菌体中检出 rll+噬菌体,所以可检出两种 不同的 rll 突变型之间重组频率极低的重组子。 Benzer 以 T4 噬菌体为材料进行了研究工作 ,正式提出“顺反子”这个术语 该方法称重组测验(recombination test),以遗传图方式确定突变子间的关 系。该法测定重组频率极灵敏,即使在 106 r II 噬菌体中只出现一个 rII+ 重组子,也可通过感染 E.coli K(λ)菌株的平板检查出来. 7.3 互补测验 7.3.1 互补测验原理和方法

基础遗传学研究首先须有突变型,然后分析突变型间的关系。重组 测验与互补测验是确定这种关系的两个基本方法」 重组测验以遗传图距确定突变的空间关系,而互补测验则是确定突变 的功能 关系。 如1区有3000多个突变型都有相同表型,这是由于所有的rⅡ突变都导 致丧失合成Ec0ⅱK(入)发育所需要一种或几种蛋白质能力,这些突变型对 E.cK(入)寄主细胞致死,但可在E,cohB菌株细胞中增殖」 既然它们有相同表型,是否它们都影响同 一种遗传功能?中这 3000多个突变型是属于 一个基因还是属于几个基因?为划分这种功能单 位界线,要讲行互补测验。 用不同l突变型成对组合同时感染大肠杆菌K(入)菌株: 如果被双重感染的细菌中产生两种亲代基因型的子代噬菌体(也有少 量重组型的噬菌体),那么必然是一个突变型补偿了另一个突变型所不具有 的功能,两个突变型称彼此互补()。如果双重感 染 的细 不产生子代噬菌体,那么这两种突变型一定有一个相同功能受到损伤。 互补测验斑点测试法(spot test) 用一种rII突变型以0.1感染比(噬菌体1细菌10)感染E.cohK(入) 菌株。噬菌体和细菌在温热的琼脂中混合,涂布在平板,琼脂凝固后在平 板上划出的 定位置上再加一滴含 一种rⅡ突变型的培养基。在这 培养基范围内, 些细菌会被两种噬菌体所感染。如在这范围内形成噬菌 斑,证明这两种突变型互补,相反不能互补。在一个培养皿平板上可做6~8 个斑点试验。 (I)rIA和rB的突变型可以互朴 rIB HIA+rIB A产物B产物 TIA rIB (2)rIB的两个突变型不能互补 rIB Q TIAt rIB A产物 +不提制 入HIA'FIB 该方法测定重组频率极敏感,重组检出率达1106,理论上可测得0.002% 的重组值,实际上所观察的最小重组频率为0.02%。根据二点杂交的结果, 可作成连锁图 互补测验结果发现,除一些缺失突变型外,突变型可分成rⅡA和「 IB两个互补群。 lA突变型的突变位点都在区的一头,是一个独立的功能单位:所 有邮突变型的突变位点都在l区的另一头,也是一个独立功能单位
基础遗传学研究首先须有突变型,然后分析突变型间的关系。重组 测验与互补测验是确定这种关系的两个基本方法。 重组测验以遗传图距确定突变的空间关系,而互补测验则是确定突变 的功能 关系。 如 rII 区有 3 000 多个突变型都有相同表型,这是由于所有的 r II 突变都导 致丧失合成 E.coli K(λ)发育所需要一种或几种蛋白质能力,这些突变型对 E.coli K(λ)寄主细胞致死,但可在 E.coli B 菌株细胞中增殖。 既然它们有相同表型,是否它们都影响同一种遗传功能?rII 中这 3 000 多个突变型是属于一个基因还是属于几个基因?为划分这种功能单 位界线,要进行互补测验。 用不同 rll 突变型成对组合同时感染大肠杆菌 K(λ)菌株: 如果被双重感染的细菌中产生两种亲代基因型的子代噬菌体(也有少 量重组型的噬菌体),那么必然是一个突变型补偿了另一个突变型所不具有 的功能,两个突变型称彼此互补(complementation)。如果双重感染的细菌 不产生子代噬菌体,那么这两种突变型一定有一个相同功能受到损伤。 互补测验斑点测试法(spot test) 用一种 r II 突变型以 0.1 感染比(噬菌体 1/细菌 10)感染 E.coli K(λ) 菌株。噬菌体和细菌在温热的琼脂中混合,涂布在平板,琼脂凝固后在平 板上划出的一定位置上再加一滴含另一种 r II 突变型的培养基。在这一滴 培养基范围内,一些细菌会被两种噬菌体所感染。如在这范围内形成噬菌 斑,证明这两种突变型互补,相反不能互补。在一个培养皿平板上可做 6~8 个斑点试验。 该方法测定重组频率极敏感,重组检出率达 1/106,理论上可测得 0.002% 的重组值,实际上所观察的最小重组频率为 0.02%。根据二点杂交的结果, 可作成连锁图 。 互补测验结果发现,除一些缺失突变型外, rII 突变型可分成 r IIA 和 r IIB 两个互补群。 rllA 突变型的突变位点都在 rll 区的一头,是一个独立的功能单位;所 有 rllB 突变型的突变位点都在 rll 区的另一头,也是一个独立功能单位