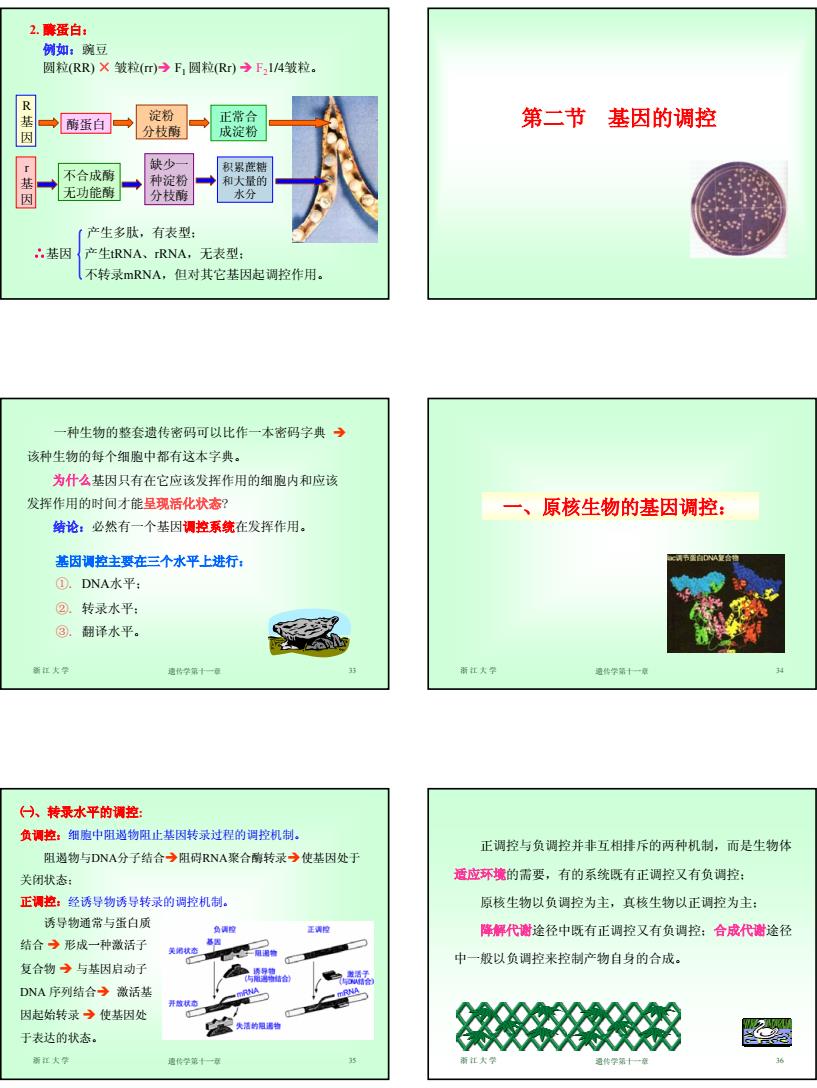
6 2. 酶蛋白: 例如:豌豆 圆粒(RR) × 皱粒(rr)Î F1 圆粒(Rr) Î F21/4皱粒。 R 基 因 酶蛋白 淀粉 分枝酶 正常合 成淀粉 r 基 因 不合成酶 无功能酶 缺少一 种淀粉 分枝酶 积累蔗糖 和大量的 水分 产生多肽,有表型; ∴基因 产生tRNA、rRNA,无表型; 不转录mRNA,但对其它基因起调控作用。 第二节 基因的调控 浙江大学 遗传学第十一章 33 一种生物的整套遗传密码可以比作一本密码字典 Î 该种生物的每个细胞中都有这本字典。 为什么基因只有在它应该发挥作用的细胞内和应该 发挥作用的时间才能呈现活化状态? 结论:必然有一个基因调控系统在发挥作用。 基因调控主要在三个水平上进行: ①. DNA水平; ②. 转录水平; ③. 翻译水平。 浙江大学 遗传学第十一章 34 一、原核生物的基因调控: 浙江大学 遗传学第十一章 35 ㈠、转录水平的调控: 负调控:细胞中阻遏物阻止基因转录过程的调控机制。 阻遏物与DNA分子结合Î阻碍RNA聚合酶转录Î使基因处于 关闭状态; 正调控:经诱导物诱导转录的调控机制。 诱导物通常与蛋白质 结合 Î 形成一种激活子 复合物 Î 与基因启动子 DNA 序列结合Î 激活基 因起始转录 Î 使基因处 于表达的状态。 浙江大学 遗传学第十一章 36 正调控与负调控并非互相排斥的两种机制,而是生物体 适应环境的需要,有的系统既有正调控又有负调控; 原核生物以负调控为主,真核生物以正调控为主; 降解代谢途径中既有正调控又有负调控;合成代谢途径 中一般以负调控来控制产物自身的合成
6 2. 酶蛋白: 例如:豌豆 圆粒(RR) × 皱粒(rr)Î F1 圆粒(Rr) Î F21/4皱粒。 R 基 因 酶蛋白 淀粉 分枝酶 正常合 成淀粉 r 基 因 不合成酶 无功能酶 缺少一 种淀粉 分枝酶 积累蔗糖 和大量的 水分 产生多肽,有表型; ∴基因 产生tRNA、rRNA,无表型; 不转录mRNA,但对其它基因起调控作用。 第二节 基因的调控 浙江大学 遗传学第十一章 33 一种生物的整套遗传密码可以比作一本密码字典 Î 该种生物的每个细胞中都有这本字典。 为什么基因只有在它应该发挥作用的细胞内和应该 发挥作用的时间才能呈现活化状态? 结论:必然有一个基因调控系统在发挥作用。 基因调控主要在三个水平上进行: ①. DNA水平; ②. 转录水平; ③. 翻译水平。 浙江大学 遗传学第十一章 34 一、原核生物的基因调控: 浙江大学 遗传学第十一章 35 ㈠、转录水平的调控: 负调控:细胞中阻遏物阻止基因转录过程的调控机制。 阻遏物与DNA分子结合Î阻碍RNA聚合酶转录Î使基因处于 关闭状态; 正调控:经诱导物诱导转录的调控机制。 诱导物通常与蛋白质 结合 Î 形成一种激活子 复合物 Î 与基因启动子 DNA 序列结合Î 激活基 因起始转录 Î 使基因处 于表达的状态。 浙江大学 遗传学第十一章 36 正调控与负调控并非互相排斥的两种机制,而是生物体 适应环境的需要,有的系统既有正调控又有负调控; 原核生物以负调控为主,真核生物以正调控为主; 降解代谢途径中既有正调控又有负调控;合成代谢途径 中一般以负调控来控制产物自身的合成
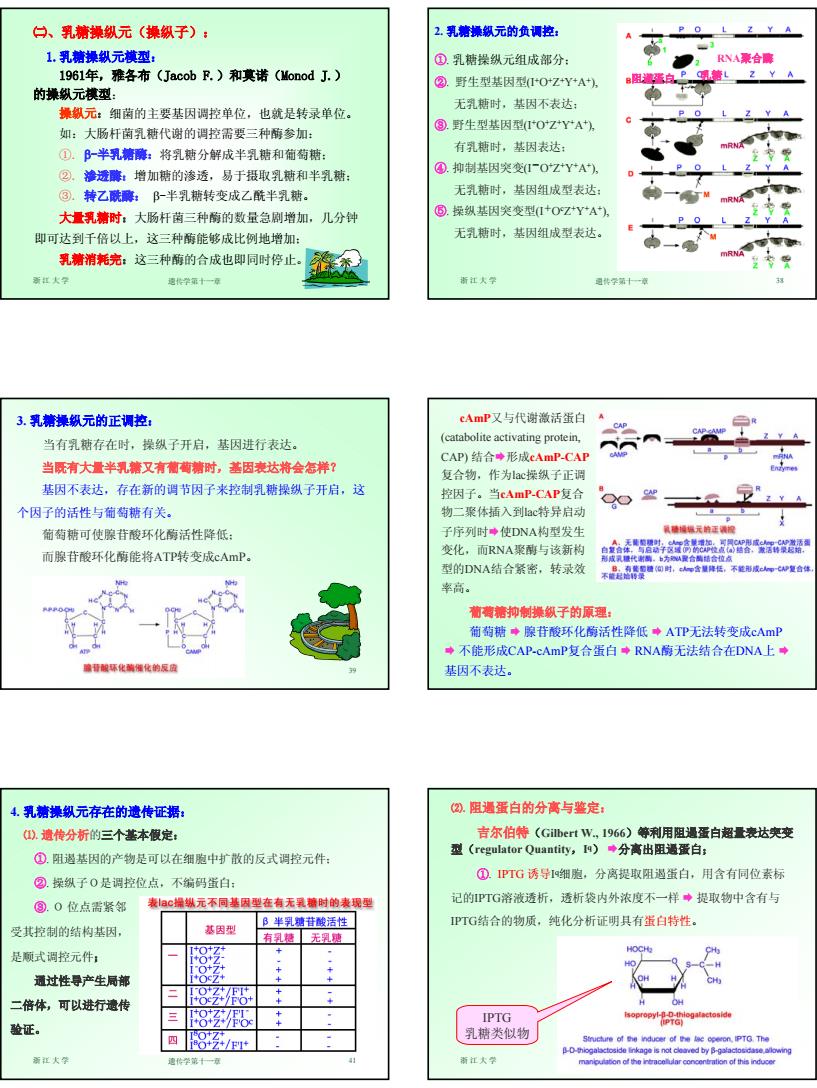
7 浙江大学 遗传学第十一章 37 ㈡、乳糖操纵元(操纵子): 1.乳糖操纵元模型: 1961年,雅各布(Jacob F.)和莫诺(Monod J.) 的操纵元模型: 操纵元:细菌的主要基因调控单位,也就是转录单位。 如:大肠杆菌乳糖代谢的调控需要三种酶参加: ①. β-半乳糖酶:将乳糖分解成半乳糖和葡萄糖; ②. 渗透酶:增加糖的渗透,易于摄取乳糖和半乳糖; ③. 转乙酰酶: β-半乳糖转变成乙酰半乳糖。 大量乳糖时:大肠杆菌三种酶的数量急剧增加,几分钟 即可达到千倍以上,这三种酶能够成比例地增加; 乳糖消耗完:这三种酶的合成也即同时停止。 浙江大学 遗传学第十一章 38 2. 乳糖操纵元的负调控: ①. 乳糖操纵元组成部分; ②. 野生型基因型(I+O+Z+Y+A+), 无乳糖时,基因不表达; ③. 野生型基因型(I+O+Z+Y+A+), 有乳糖时,基因表达; ④. 抑制基因突变(I-O+Z+Y+A+), 无乳糖时,基因组成型表达; ⑤. 操纵基因突变型(I+Oc Z+Y+A+), 无乳糖时,基因组成型表达。 阻遏蛋白 乳糖 RNA聚合酶 浙江大学 遗传学第十一章 39 3. 乳糖操纵元的正调控: 当有乳糖存在时,操纵子开启,基因进行表达。 当既有大量半乳糖又有葡萄糖时,基因表达将会怎样? 基因不表达,存在新的调节因子来控制乳糖操纵子开启,这 个因子的活性与葡萄糖有关。 葡萄糖可使腺苷酸环化酶活性降低; 而腺苷酸环化酶能将ATP转变成cAmP。 cAmP又与代谢激活蛋白 (catabolite activating protein, CAP) 结合¨形成cAmP-CAP 复合物,作为lac操纵子正调 控因子。当cAmP-CAP复合 物二聚体插入到lac特异启动 子序列时¨使DNA构型发生 变化,而RNA聚酶与该新构 型的DNA结合紧密,转录效 率高。 葡萄糖抑制操纵子的原理: 葡萄糖 ¨ 腺苷酸环化酶活性降低 ¨ ATP无法转变成cAmP ¨ 不能形成CAP-cAmP复合蛋白 ¨ RNA酶无法结合在DNA上 ¨ 基因不表达。 浙江大学 遗传学第十一章 41 4. 乳糖操纵元存在的遗传证据: ⑴. 遗传分析的三个基本假定: ①. 阻遏基因的产物是可以在细胞中扩散的反式调控元件; ②. 操纵子O是调控位点,不编码蛋白; ③. O 位点需紧邻 受其控制的结构基因, 是顺式调控元件; 通过性导产生局部 二倍体,可以进行遗传 验证。 浙江大学 遗传学第十一章 42 ⑵. 阻遏蛋白的分离与鉴定: 吉尔伯特(Gilbert W., 1966)等利用阻遏蛋白超量表达突变 型(regulator Quantity,Iq) ¨分离出阻遏蛋白; ①. IPTG 诱导Iq细胞,分离提取阻遏蛋白,用含有同位素标 记的IPTG溶液透析,透析袋内外浓度不一样 ¨ 提取物中含有与 IPTG结合的物质,纯化分析证明具有蛋白特性。 IPTG 乳糖类似物
7 浙江大学 遗传学第十一章 37 ㈡、乳糖操纵元(操纵子): 1.乳糖操纵元模型: 1961年,雅各布(Jacob F.)和莫诺(Monod J.) 的操纵元模型: 操纵元:细菌的主要基因调控单位,也就是转录单位。 如:大肠杆菌乳糖代谢的调控需要三种酶参加: ①. β-半乳糖酶:将乳糖分解成半乳糖和葡萄糖; ②. 渗透酶:增加糖的渗透,易于摄取乳糖和半乳糖; ③. 转乙酰酶: β-半乳糖转变成乙酰半乳糖。 大量乳糖时:大肠杆菌三种酶的数量急剧增加,几分钟 即可达到千倍以上,这三种酶能够成比例地增加; 乳糖消耗完:这三种酶的合成也即同时停止。 浙江大学 遗传学第十一章 38 2. 乳糖操纵元的负调控: ①. 乳糖操纵元组成部分; ②. 野生型基因型(I+O+Z+Y+A+), 无乳糖时,基因不表达; ③. 野生型基因型(I+O+Z+Y+A+), 有乳糖时,基因表达; ④. 抑制基因突变(I-O+Z+Y+A+), 无乳糖时,基因组成型表达; ⑤. 操纵基因突变型(I+Oc Z+Y+A+), 无乳糖时,基因组成型表达。 阻遏蛋白 乳糖 RNA聚合酶 浙江大学 遗传学第十一章 39 3. 乳糖操纵元的正调控: 当有乳糖存在时,操纵子开启,基因进行表达。 当既有大量半乳糖又有葡萄糖时,基因表达将会怎样? 基因不表达,存在新的调节因子来控制乳糖操纵子开启,这 个因子的活性与葡萄糖有关。 葡萄糖可使腺苷酸环化酶活性降低; 而腺苷酸环化酶能将ATP转变成cAmP。 cAmP又与代谢激活蛋白 (catabolite activating protein, CAP) 结合¨形成cAmP-CAP 复合物,作为lac操纵子正调 控因子。当cAmP-CAP复合 物二聚体插入到lac特异启动 子序列时¨使DNA构型发生 变化,而RNA聚酶与该新构 型的DNA结合紧密,转录效 率高。 葡萄糖抑制操纵子的原理: 葡萄糖 ¨ 腺苷酸环化酶活性降低 ¨ ATP无法转变成cAmP ¨ 不能形成CAP-cAmP复合蛋白 ¨ RNA酶无法结合在DNA上 ¨ 基因不表达。 浙江大学 遗传学第十一章 41 4. 乳糖操纵元存在的遗传证据: ⑴. 遗传分析的三个基本假定: ①. 阻遏基因的产物是可以在细胞中扩散的反式调控元件; ②. 操纵子O是调控位点,不编码蛋白; ③. O 位点需紧邻 受其控制的结构基因, 是顺式调控元件; 通过性导产生局部 二倍体,可以进行遗传 验证。 浙江大学 遗传学第十一章 42 ⑵. 阻遏蛋白的分离与鉴定: 吉尔伯特(Gilbert W., 1966)等利用阻遏蛋白超量表达突变 型(regulator Quantity,Iq) ¨分离出阻遏蛋白; ①. IPTG 诱导Iq细胞,分离提取阻遏蛋白,用含有同位素标 记的IPTG溶液透析,透析袋内外浓度不一样 ¨ 提取物中含有与 IPTG结合的物质,纯化分析证明具有蛋白特性。 IPTG 乳糖类似物