
(al oma,a cancer of the retina. b RB位于13q14,编码视网膜母细胞瘤蛋白Rb,能结合肿瘤抗原,(SV40T抗 原)调控细胞周期,抑制细胞增殖,最早在视网膜母细胞瘤的研究中被发现。 na2e omc tumor fomm ation Kundson早在1971年提出著名的“二次突变假设(two mutation hypothsis).”。后来遗传学家发现家族性患儿的体细胞中13号染色体存在缺失 (13q14),表明该区域可能存在与视网膜母细胞瘤发生有关的基因Rb。 Sparkes(1980)将Rb基因定位于13q14。Benedict(1983)提出位于13ql4的Rb 的一对等位基因均失活才会产生该肿瘤。表明Rb是以隐性方式起作用。 Stephe等(1986)开始构建了人的13号染色体的λ噬菌体文库,其中1.5Kb的 DNA片段和Rb有关。接若又分离到一个3OKb的cDNA片段,用此片段可检测 到1314带70Kb区域的基因。同年眼科专家】 y 发Rh生活时可可南 杂合子(Rb+Rb)易被诱变而失活:他还发现经眼球摘除手术能存活下来的患 儿长大后易患成骨肉嘉,乳腺癌和小细胞肺癌。 1986年Friend将13q14位点的Rb经cDNA克隆制成各种探针,检查很多例
RB 位于 13q14,编码视网膜母细胞瘤蛋白 Rb,能结合肿瘤抗原,(SV40 T 抗 原)调控细胞周期, 抑制细胞增殖,最早在视网膜母细胞瘤的研究中被发现。 Kundson 早在 1971 年提出著名的 “ 二次突变假设(two mutation hypothsis)”。后来遗传学家发现家族性患儿的体细胞中 13 号染色体存在缺失 (13q14),表明该区域可能存在与视网膜母细胞瘤发生有关的基因 Rb。 Sparkes(1980)将 Rb 基因定位于 13q14。Benedict(1983)提出位于 13q14 的 Rb 的一对等位基因均失活才会产生该肿瘤。表明 Rb 是以隐性方式起作用。 Stephe 等(1986)开始构建了人的 13 号染色体的λ噬菌体文库,其中 1.5Kb 的 DNA 片段和 Rb 有关。接着又分离到一个 30Kb 的 cDNA 片段,用此片段可检测 到 13q14 带 70Kb 区域的基因。同年眼科专家 Dryja 发现 Rb 失活时可以致瘤, 杂合子(Rb+/Rb-)易被诱变而失活; 他还发现经眼球摘除手术能存活下来的患 儿长大后易患成骨肉瘤,乳腺癌和小细胞肺癌。 1986 年 Friend 将 13q14 位点的 Rb 经 cDNA 克隆制成各种探针,检查很多例

骨肉瘤和组织肉瘤患者发现约50%的患者贴基因纯合缺失。 跑中, 培养的 伟目 逆转。更进一步证明了R贴的抑癌作用。他克隆了b全长的cDA,发现其 物是一种长928氨基酸,分子量为105Kda的蛋白质。 二、重复 (一)重复的类型 1203456 L20343456 顺接重复 L20344356 反接重复 L34,2.3456 异臂重复 1234256 移位重复 图22.重复的类型 (二)重复产生的原因: 1.断裂-融合桥的形成: 23 2.染色体纽结: 3.不等交换: 4.非同源性重组 23 5转座效应 24 重复的细胞学鉴定: 细胞学特征 同源染色体联会时可见: 重复环 染色体末端不配对而突出 注意: 风分重复环与缺失环 当重复区段很短时很难观察到重复环 面复纯合体也视空不到面复不 (三)重复的遗传效应: 1.主要通过雌配子遗传. 2.位置效应(position effect) 3.剂量效应(position effect)
骨肉瘤和组织肉瘤患者发现约 50%的患者 Rb 基因纯合缺失。 1987 年 Stanbridge 等分离了人类 13 号染色体,将它注入培养的成骨肉瘤细 胞中,结果导致了癌细胞逆转,同年李文华把 Rb 转化到培养的癌细胞中使其 逆转。更进一步证明了 Rb 的抑癌作用。他克隆了 Rb 全长的 cDNA,发现其产 物是一种长 928 氨基酸,分子量为 105Kda 的蛋白质。 二、重复 (一) 重复的类型: (二)重复产生的原因: 1.断裂-融合桥的形成: 2.染色体纽结: 3.不等交换: 4.非同源性重组 5.转座效应 重复的细胞学鉴定: 细胞学特征 同源染色体联会时可见: 重复环 染色体末端不配对而突出 注意: 区分重复环与缺失环 当重复区段很短时很难观察到重复环 重复纯合体也观察不到重复环 (三)重复的遗传效应: 1.主要通过雌配子遗传. 2.位置效应(position effect) 3.剂量效应(position effect). 1 2 3 4 5 6 1 2 3 4 3 4 5 6 顺接重复 1 2 3 4 4 3 5 6 反接重复 1 3 4 2 3 4 5 6 异臂重复 1 2 3 4 a b 5 6 移位重复 图 22- 重复的类型 1 2 3 4 1 2 3 4 1 2 3 3 4 1 2 4

Duplications A B C D A@D ABC A B C D AD E A B C D Duplication loop A R Asymmetrical pairing eye v+red vs v vermei v+v red v+vv vermeil 51 a Position effeet
Duplications Duplication loop Asymmetrical pairing Dosage effect eg. eye color of Drosophila v+ red vs v vermeil v+v red v+vv vermeil Position effect
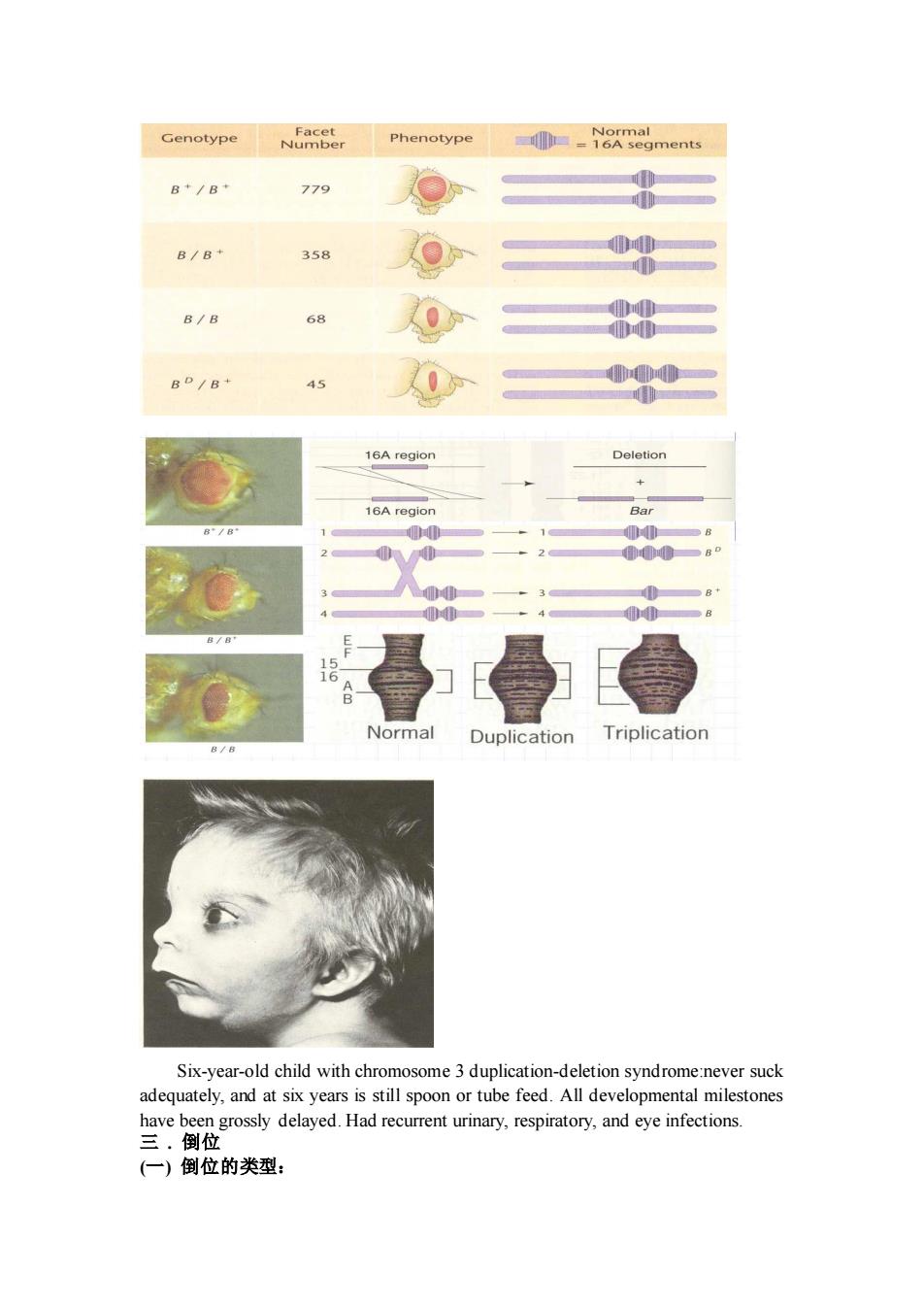
Genotype Phenotype -1oA segments B+/B+ 779 B/B+ 358 8/8 0分 8D/8- 45 0 16A region Deletion 16A reglon Normal Duplicatic Triplication Six-year-old child with chromosome3 duplication-deletion syndrome:never suck adequately,and at six years is still spoon or tube feed.All developmental milestones have been grossly delayed.Had recurrent urinary,respiratory,and eye infections. 三.倒位 (一)倒位的类型:
Six-year-old child with chromosome 3 duplication-deletion syndrome:never suck adequately, and at six years is still spoon or tube feed. All developmental milestones have been grossly delayed. Had recurrent urinary, respiratory, and eye infections. 三 . 倒位 (一) 倒位的类型:

臂内倒位(pracentric inversion) 臂间倒位(pericentric inversion) (仁)倒位的产生的原因 染色体纽结、断裂和重接 (②)转座因子可引起染色体倒位 (3)自发倒位:沙门氏细菌的相转变Mu噬菌体的G片段的倒位 2兰 不画 123 123 图222染色体纽结产生倒位 倒位的细胞学鉴定 细胞学特征 同源染色体联会时: 倒位区段过长 —倒过来配对,其余游离倒位区段较短一正常部分配对, 其余不配对 倒位区段适中—形成倒位圈 注意 区分倒位圈与缺失、重复圈的结构差异倒位纯合体无明显细胞学特征 倒位杂合体的联会 倒位杂合体及其联会 c d e f b c d e f 倒位区段过长的 倒位杂合体之联会 (三)倒位遗传效应: 1降低了倒位杂合体的连锁基因重组率。 2.引起基因重排
臂内倒位(pracentric inversion) 臂间倒位(pericentric inversion) (二) 倒位的产生的原因: (1) 染色体纽结、断裂和重接 (2) 转座因子可引起染色体倒位 (3) 自发倒位:沙门氏细菌的相转变 Mu 噬菌体的 G 片段的倒位 倒位的细胞学鉴定 细胞学特征 同源染色体联会时: 倒位区段过长——倒过来配对,其余游离倒位区段较短——正常部分配对, 其余不配对 倒位区段适中——形成倒位圈 注意 区分倒位圈与缺失、重复圈的结构差异倒位纯合体无明显细胞学特征 倒位杂合体的联会 (三)倒位遗传效应: 1.降低了倒位杂合体的连锁基因重组率。 2.引起基因重排。 1 2 3 4 1 2 3 4 1 2 3 4 纽结 1 断裂重接 2 4 3 1 2 3 4 1 2 3 4 4 2 4 2 3 3 图 22-2 染色体纽结产生倒位