
-35 -10 +1 5TTGACA- TATAATStart site The first nucleotide (the start site)of a transcribed DNA sequence is denoted as +1 and the second one as +2;the ● nucleotide preceding the start site is denoted as-1.These designations refer to the coding strand of DNA.Recall that the ● sequence of the template strand ofDNA is the complement of that of the RNA transcript (see Figure 5.26).In contrast, the coding strand of DNA has the same sequence as that of the RNA transcript except for thymine (T)in place of uracil (U).The coding strand is also known as the sense (+ strand,and the template strand as the antisense (-)strand
• The first nucleotide (the start site) of a transcribed DNA sequence is denoted as +1 and the second one as +2; the • nucleotide preceding the start site is denoted as -1. These designations refer to the coding strand of DNA. Recall that the • sequence of the template strand of DNA is the complement of that of the RNA transcript (see Figure 5.26). In contrast, • the coding strand of DNA has the same sequence as that of the RNA transcript except for thymine (T) in place of uracil • (U). The coding strand is also known as the sense (+) strand, and the template strand as the antisense (-) strand

启动子 在RNA合成中用作模板的链称模板链或负 (-)链。与模板链互补的DNA链称为非模板 链或正(+)链。非模板链与转录的RNA在碱 基序列上是一致的,只是DNA中用T代替了U 。非模板链有时又称为编码链,它在转录和蛋 白质合成中没有直接功能。RNA合成中转录起 始点标上+1,这个位点前面的碱基对标以负数 ,它们是不被转录的,而起始之后的碱基序列 标以正数。这里不设0数。5'端方向称上游(up stream)3'端方向称下游(downstream)根据习 惯是指非模板链中的序列
启动子 在RNA合成中用作模板的链称模板链或负 (-)链。与模板链互补的DNA链称为非模板 链或正(+)链。非模板链与转录的RNA在碱 基序列上是一致的,只是DNA中用T代替了U 。非模板链有时又称为编码链,它在转录和蛋 白质合成中没有直接功能。RNA合成中转录起 始点标上+1,这个位点前面的碱基对标以负数 ,它们是不被转录的,而起始之后的碱基序列 标以正数。这里不设0数。5΄端方向称上游(up stream)3΄端方向称下游(downstream)根据习 惯是指非模板链中的序列

·实验证明,启动子序列中某些核苷酸是保守的 ,即在大多数情况下它们都会出现,称为共有 序列。原核与真核生物启动子通常含有两个共 有序列。原核生物共有序列出现在-10和-35所 以称-10区和-35区。真核生物共有序列出现在 -25和-75,分别称为-25区(TATA box:或 Hoghess box)和-75区(CAAT box)。紧 邻转录起始点的共有序列是一个A/T富含区, 它使DNA双链容易局部解链
• 实验证明,启动子序列中某些核苷酸是保守的 ,即在大多数情况下它们都会出现,称为共有 序列。原核与真核生物启动子通常含有两个共 有序列。原核生物共有序列出现在-10和-35所 以称-10区和-35区。真核生物共有序列出现在 -25和-75,分别称为-25区(TATA box或 Hoghess box)和-75区(CAAT box)。紧 邻转录起始点的共有序列是一个A/T富含区, 它使DNA双链容易局部解链
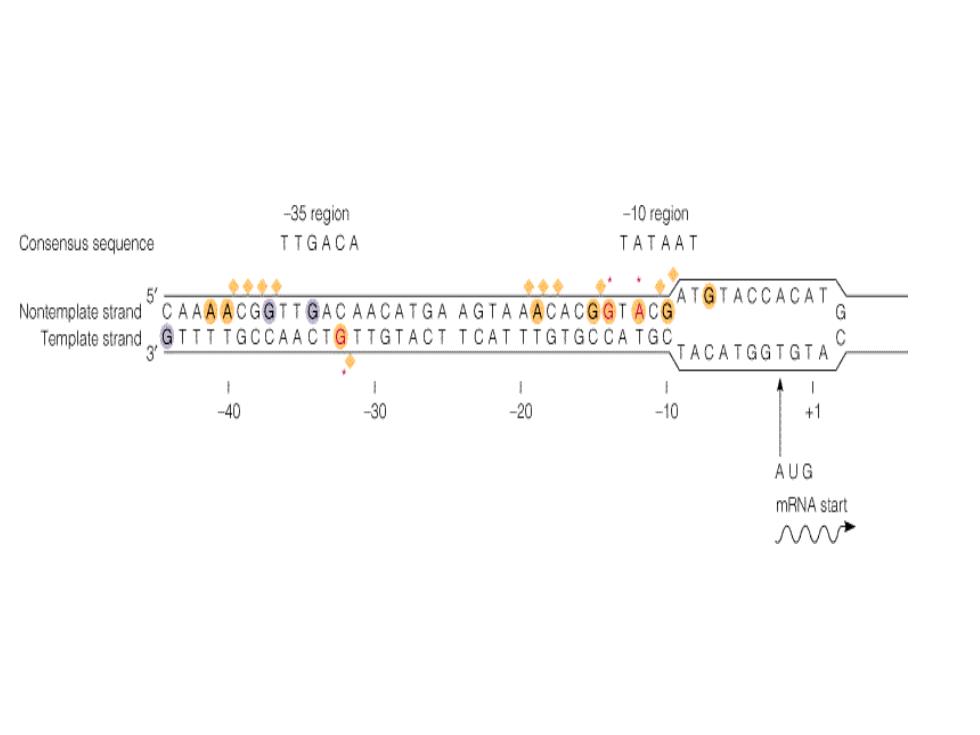
-35 region -10 region Consensus sequence TTGACA TATAAT 5 ●◆女 ◆◆”·◆/ATGTACCACAT Nontemplate strand CAAAACGGTTGAC AACATGA AGT A AACACGGT ACG Template strand。 GTTT TGCCAACTGTTGTACT TCAT TTGTGCCATGC C TACATGGTGTA 1 -40 30 -20 -10 AUG mRNA start 产

五起始 1.6亚基与核心酶结合形成RNA聚合酶全酶。 2.全酶以静电作用和启动子上游非特异性DNA结合。 3. RNA聚合酶全酶沿DNA上下游动,包括共有序列在内 与启动子特异性结合, 全酶结合后引起DNA片段部分解 链伸展开约11bp(从-9~+2) 4. 酶与ATP或GTP结合,结合的三磷酸核苷酸形成转录 物的5'末端,刚结合上的三磷酸核苷3 OH与下一个 核苷酸生成磷酸酯键。所以真核和原核生物的转录作用 以A或G开始。 5.当全酶继续向下游移动,约有4个核苷酸与ATP(GTP 进行聚合。δ亚基从全酶上解离,核心酶继续合成。 新RNA链生长方向是5'3',新合成的RNA链与被转 录的DNA链互补,反平行。 Sigma Subunits of RNA Polymerase Recognize Promoter Sites
五 起 始 1.δ亚基与核心酶结合形成RNA聚合酶全酶。 2.全酶以静电作用和启动子上游非特异性DNA 结合。 3.RNA聚合酶全酶沿DNA上下游动,包括共有序列在内 与启动子特异性结合,全酶结合后引起DNA片段部分解 链伸展开约11bp(从-9~+2) 4.酶与ATP或GTP结合,结合的三磷酸核苷酸形成转录 物的5΄末端,刚结合上的三磷酸核苷3΄→OH与下一个 核苷酸生成磷酸酯键。所以真核和原核生物的转录作用 以A或G开始。 5.当全酶继续向下游移动,约有4个核苷酸与ATP(GTP )进行聚合。δ亚基从全酶上解离,核心酶继续合成。 新RNA链生长方向是5΄→3΄,新合成的RNA链与被转 录的DNA链互补,反平行。 Sigma Subunits of RNA Polymerase Recognize Promoter Sites