
昆虫知识ENTOMOLOGICALKNOWLEDGE2003.40(3).280..基础知识昆虫的肢体自残现象王孟卿彩万志“*(中国农业大学昆虫系北京100094)Autotomy in insects. WANGMeng Qing, CAI War Zhi(Departagit g Entomology,Ciner Agacultu al Lir'ersity ,Bejing100094,China)AbstractThetypes,advantages,disadvantages,causes anc ccokgical importanceof autotomy in insects,and the taxain which it occurs ,are summeri zed.Key wordsinsects,autotomy,behavior,antrpredator defence,evolution摘要该文主要就昆虫的肢体自残现象发生的种类和虫态、被残器官和部位、自残发生的原因、自残对昆虫个体的有利及不利影响进行了综述,并简单地讨论了肢体自残现象在进化上的意义。关键词昆虫,肢体自残,行为,自卫,进化肢体自残(autotomy)的概念由Fredericq1发生肢体自残的昆虫类群、虫态和被残器官1883年首次提出,其定义尚未统一。本文中1.1发生肢体自残的昆虫类群的体自残是指当某种动物的某一个体遭到攻击(或遇到其它不利条件)时其部分肢体会很快已发现能够肢体自残的昆虫见于双尾目、目、蜻蜓目、等翅目、螳螂目、目、直翅目、脱落从而使该个体得以生存的生物学现象,是动物在被动情况下个体水平上的一种应急对同翅目、半翅目、鳞翅目、膜翅目、鞘翅目、双翅目等类群[4~25],即在无翅亚纲和有翅亚纲或表策。因此像一些社会性昆虫的生殖型雌性个体(如重后)在交配后为繁殖后代之便而自动断变态、原变态、不全变态和全变态类的昆虫中都翅的现象不属于自残范畴:蜜蜂在自卫董人或有肢体自残的种类,尤以直翅类昆虫最为普遍其它动物时毒针等器官留在被董对象体内,虽但并非在这些目中所有的种类都具有肢体自残然蜜蜂在种群水平受到保护,但董人的蜜蜂个习性。1.2自残发生的虫态体却因器官缺损而亡,显然,按本文中肢体自残的概念蜜蜂董人不应该属于肢体自残现象,而具有肢体自残习性的昆虫并非各个虫态都是社会性昆虫的自卫方式之一。另外,值得注能发生肢体自残,一般不全变态的种类(如直翅意的是中文文献中有时把同种或同类动物不同类昆虫)若虫期和成虫期均能发生自残,而全变个体间的自相残杀现象(cannibalism)简称自残,态类的昆虫大多发生在成虫期。这与肢体自残是2个截然不同的概念。肢体自1.3被残器官残现象在动物界普遍存在[2,3],如壁虎断尾和螃从自残的概念分析自残器官应是虫体的次蟹断肢等。其实,肢体自残现象在昆虫纲中也要部分实际情况也正是如此。从以前的记载常有发生但有关的报道比较零散。在此作者·国家自然科学基金资助项目(No.30070518)。仅就昆虫的肢体自残现象加以介绍。**通讯作者,Email:caiwz@cau.edu.cn收稿日期:2002-08-11,修回日期:2002-11-20D1994-2007ChinaAcademicJournal ElectronicPublishingHouse.All rightsreservedhtp:/hnr.cnki.ner
基础知识 昆虫的肢体自残现象3 王孟卿 彩万志33 (中国农业大学昆虫系 北京 100094) Autotomy in insects. WANG Meng2Qing , CAI Wan2Zhi ( Department of Entomology , China Agricultural University , Beijing 100094 ,China) . Abstract The types ,advantages ,disadvantages ,causes and ecological importance of autotomy in insects ,and the taxa in which it occurs ,are summarized. Key words insects , autotomy , behavior , anti2predator defence , evolution 摘 要 该文主要就昆虫的肢体自残现象发生的种类和虫态、被残器官和部位、自残发生的原因、自残 对昆虫个体的有利及不利影响进行了综述 ,并简单地讨论了肢体自残现象在进化上的意义。 关键词 昆虫 , 肢体自残 , 行为 , 自卫 , 进化 3 国家自然科学基金资助项目(No. 30070518) 。 33 通讯作者 ,E2mail :caiwz @cau. edu. cn 收稿日期 :2002208211 ,修回日期 :2002211220 肢体自残 ( autotomy) 的概念由 Frédéricq 1883 年首次提出[1 ] ,其定义尚未统一。本文中 的肢体自残是指当某种动物的某一个体遭到攻 击(或遇到其它不利条件) 时其部分肢体会很快 脱落从而使该个体得以生存的生物学现象 ,是 动物在被动情况下个体水平上的一种应急对 策。因此 ,像一些社会性昆虫的生殖型雌性个 体(如 后) 在交配后为繁殖后代之便而自动断 翅的现象不属于自残范畴 ;蜜蜂在自卫蜇人或 其它动物时毒针等器官留在被蜇对象体内 ,虽 然蜜蜂在种群水平受到保护 ,但蜇人的蜜蜂个 体却因器官缺损而亡 ,显然 ,按本文中肢体自残 的概念蜜蜂蜇人不应该属于肢体自残现象 ,而 是社会性昆虫的自卫方式之一。另外 ,值得注 意的是中文文献中有时把同种或同类动物不同 个体间的自相残杀现象(cannibalism) 简称自残 , 这与肢体自残是 2 个截然不同的概念。肢体自 残现象在动物界普遍存在[2 ,3 ] ,如壁虎断尾和螃 蟹断肢等。其实 ,肢体自残现象在昆虫纲中也 常有发生 ,但有关的报道比较零散。在此 ,作者 仅就昆虫的肢体自残现象加以介绍。 1 发生肢体自残的昆虫类群、虫态和被残器官 111 发生肢体自残的昆虫类群 已发现能够肢体自残的昆虫见于双尾目、 蜉蝣目、蜻蜓目、等翅目、螳螂目、 目、直翅目、 同翅目、半翅目、鳞翅目、膜翅目、鞘翅目、双翅 目等类群[4~25 ] ,即在无翅亚纲和有翅亚纲或表 变态、原变态、不全变态和全变态类的昆虫中都 有肢体自残的种类 ,尤以直翅类昆虫最为普遍 , 但并非在这些目中所有的种类都具有肢体自残 习性。 112 自残发生的虫态 具有肢体自残习性的昆虫并非各个虫态都 能发生肢体自残 ,一般不全变态的种类(如直翅 类昆虫) 若虫期和成虫期均能发生自残 ,而全变 态类的昆虫大多发生在成虫期。 113 被残器官 从自残的概念分析自残器官应是虫体的次 要部分 ,实际情况也正是如此。从以前的记载 · 082 · 昆虫知识 ENTOMOLOGICAL KNOWLEDGE 2003 40(3) © 1994-2007 China Academic Journal Electronic Publishing House. All rights reserved. http://www.cnki.net
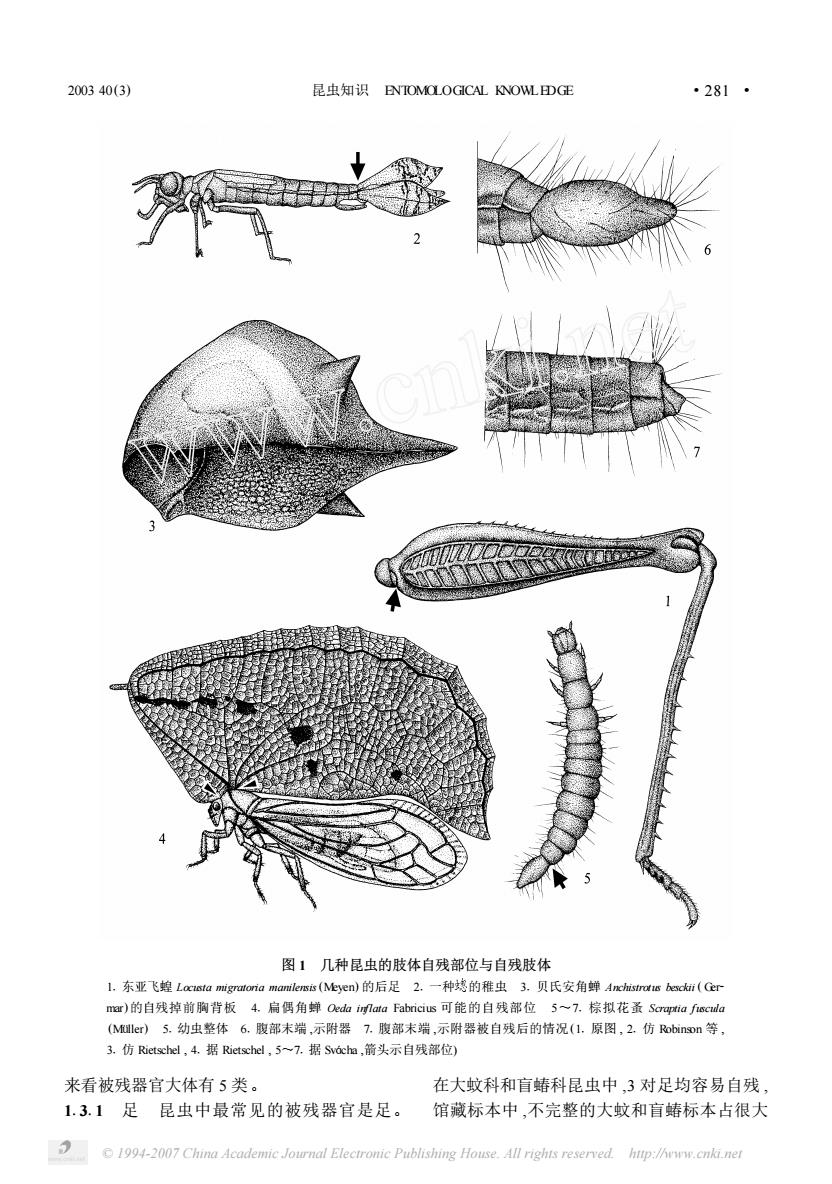
2003 40(3)昆虫知识ENTOMOLOGICALKNOWLEDGE.281:图1:几种昆虫的肢体自残部位与自残肢体1.东亚飞蝗Locistamigratoriamamilensis(Meyen)的后足2.一种媳的稚虫3.贝氏安角蝉Anchistrotisbescki(Germar)的自残掉前胸背板4.扁偶角蝉OedainflataFabricius可能的自残部位5~7.棕拟花蛋Scraptiafiscudla(Muller)5.幼虫整体6.腹部末端,示附器7.腹部末端,示附器被自残后的情况(1.原图,2.仿Robinson等,3.仿Rietschel,4.据Rietschel,57.据Svocha,箭头示自残部位)来看被残器官大体有5类。在大蚊科和盲蝽科昆虫中3对足均容易自残1.3.1足昆虫中最常见的被残器官是足。馆藏标本中,不完整的大蚊和盲蝽标本占很大1994-2007ChinaAcademicJournal ElectronicPublishingHouse.All righisreservedhup:/hemw.cnki.ner
图 1 几种昆虫的肢体自残部位与自残肢体 11 东亚飞蝗 Locusta migratoria manilensis (Meyen) 的后足 21 一种 的稚虫 31 贝氏安角蝉 Anchistrotus besckii ( Ger2 mar)的自残掉前胸背板 41 扁偶角蝉 Oeda inflata Fabricius 可能的自残部位 5~71 棕拟花蚤 Scraptia fuscula (Müller) 51 幼虫整体 61 腹部末端 ,示附器 71 腹部末端 ,示附器被自残后的情况(11 原图 , 21 仿 Robinson 等 , 31 仿 Rietschel , 41 据 Rietschel , 5~71 据 Svácha ,箭头示自残部位) 来看被残器官大体有 5 类。 11311 足 昆虫中最常见的被残器官是足。 在大蚊科和盲蝽科昆虫中 ,3 对足均容易自残 , 馆藏标本中 ,不完整的大蚊和盲蝽标本占很大 2003 40(3) 昆虫知识 ENTOMOLOGICAL KNOWLEDGE · 182 · © 1994-2007 China Academic Journal Electronic Publishing House. All rights reserved. http://www.cnki.net

.282.昆虫知识ENTOMOLOGICALKNOWLEDGE2003 40(3)比例,英、美学者称这两类昆虫为自杀性昆虫部位在第8腹节和附器基部之间。在身体主体(suicideinsects)。直翅类昆虫的后足,特别是蝗和被残器官间有一层膜能有效阻止自残后血液科昆虫发达的跳跃足(图1:1)是最常见的被残外流。器官之一。2引起自残的原因1.3.2尾须双尾目昆虫的尾须在被捉时尾须常从基部断掉。缨尾目、浮目等昆虫可能引起昆虫自残的原因主要有以下几个方也有类似的情况。面。1.3.3尾鳃蜻蜓目中类昆虫稚虫腹部末2.1捕食者和人类的威胁这是昆虫自残最端有3对鳃叶状构造(图1:2),在密度较高时,主要的原因。当自残者被人类或捕食者握住或常会自残1~3个尾鳃,以减少自相残杀。住自残器官或当其受到捕食者威胁时,有自1.3.4前胸背板同翅目角蝉科昆虫以其发残能力的昆虫会很快断掉自残器官而逃跑。如达而形状奇特的前胸背板而备受鼠虫形态学家果您提到蝗科或蠡斯科等直翅目昆虫,用食指和分类学家的关注,前胸背板的功能与其在进和拇指握住其一只或一对后足,被握的足很快化上的意义更是大家探索的热点之一[26~30],一会被自残者断掉。般认为其功能主要是拟态和感觉。但是,2.2意外伤害当蟋蜂等直翅目昆虫的后足Struimpel和Rietschel的研究则认为至少部分角的一部分受到伤害(如腔节断掉)时这些直翅蝉发达的前胸背板能自残以抵御鸟类等敌害的自昆虫常常会从转节和股节的连接处把大部分进攻(图1:3,4)[12,13)。后足断掉。1.3.5腹末附器鞘翅目拟花蚤科拟花蚤属2.3接触毒物或杀虫剂首次用化学成分诱Scraptia和幻拟花蚤属Nothotelus的幼虫(可能导昆虫肢体自残的报道见于1943年,Emmel用还有其它拟花蚤科的幼虫)在腹部第8节末端DDT处理花翅按蚊AnophelesmaculipennisMeigen能使按蚊后足自残18。1944年,Wiesmann和有1个近卵形的附器(图1:5,6),Boving和Craighead曾注意到该附器容易折断,Watt曾推Fenjves报道了尖音库蚊CulexpipiensLinnaeus、测此构造可能在遇到捕食者时自残以利幼虫逃麦蛾Sitotrogacerealella(Olivier)及一种尺跑,Svacha对棕拟花蚤Scraptiafuscula(Muller)Cheimatobiabrumata(Linnaeus)接触DDT后成虫的幼虫腹部末端第9节的附器做了详细研究后足自残的现象[9。Brousse-Gaury发现用氰化江发现附器表面有细长的刚毛,刚毛作为触觉感钾注入或用甲醛熏蒸均能引起家蟋蜂的后足自受器,一旦被触及马上做出反应:幼虫的腹部向残7。Uvarov也曾提及当把蝗虫放入毒瓶时蝗虫也会发生后足自残4)。20世纪80年代以一侧弯曲,使尾部对着入侵者,如果附器被擒住,自残就会发生(图1:7):如果入侵者动作很来,Moore及其合作者对小菜蛾Plutellaxylastella(Linnaeus))【101]、Ortego和Bowers对美洲蚱轻(附肢没有变形或毁坏),甲虫则逃跑2)。并且发现经过1次蜕皮后其相应部位能够再生。Schistocercacamericana(Drury)的药剂诱导自残做了比较详细的研究[23]。1.4自残部位凡是能够肢体自残的种类均有特定的自残2.4种群密度过大对一些具有自相残杀习部位,一般位于与身体连接最少的部分。如以性的蜻蜓目稚虫而言,在密度较高时尾鳃增加足自残的昆虫自残部位为转节与股节之间,以了被同类捕食的机会,因此在实验条件下随着尾须和触自残的昆虫自残部位为尾须和鳃的基稚虫密度的增大,尾鳃自残的比例也就上升[15,16] 。部,贝氏安角蝉Anchistrotusbescki(Germar)的自残部位在前胸背板膨大部分的基部,而棕拟花此外在实验条件下,一些物理刺激(如机蚤Scraptiafuscula(Muller)的幼虫腹末附器自残械压力、温度变化和电击等)也能引起蝗虫和蟋D1994-2007ChinaAcademicJournal ElectronicPublishingHouse.All righisresenved.htp:/hn.cnki.ne
比例 ,英、美学者称这两类昆虫为自杀性昆虫 (suicide insects) 。直翅类昆虫的后足 ,特别是蝗 科昆虫发达的跳跃足(图 1 :1) 是最常见的被残 器官之一。 11312 尾须 双尾目昆虫的尾须在被捉时尾 须常从基部断掉。缨尾目、蜉蝣目等昆虫可能 也有类似的情况。 11313 尾鳃 蜻蜓目中 类昆虫稚虫腹部末 端有 3 对鳃叶状构造(图 1 :2) ,在密度较高时 , 常会自残 1~3 个尾鳃 ,以减少自相残杀。 11314 前胸背板 同翅目角蝉科昆虫以其发 达而形状奇特的前胸背板而备受昆虫形态学家 和分类学家的关注 ,前胸背板的功能与其在进 化上的意义更是大家探索的热点之一[26~30 ] ,一 般认 为 其 功 能 主 要 是 拟 态 和 感 觉。但 是 , Strümpel 和 Rietschel 的研究则认为至少部分角 蝉发达的前胸背板能自残以抵御鸟类等敌害的 进攻(图 1 :3 ,4) [12 ,13 ] 。 11315 腹末附器 鞘翅目拟花蚤科拟花蚤属 Scraptia 和幻拟花蚤属 Nothotelus 的幼虫 (可能 还有其它拟花蚤科的幼虫) 在腹部第 8 节末端 有 1 个近卵形的附器 (图 1 : 5 , 6) , Boving 和 Craighead 曾注意到该附器容易折断 ,Watt 曾推 测此构造可能在遇到捕食者时自残以利幼虫逃 跑 ,Svácha 对棕拟花蚤 Scraptia fuscula (Müller) 的幼虫腹部末端第 9 节的附器做了详细研究 , 发现附器表面有细长的刚毛 ,刚毛作为触觉感 受器 ,一旦被触及马上做出反应 :幼虫的腹部向 一侧弯曲 ,使尾部对着入侵者 ,如果附器被擒 住 ,自残就会发生(图 1 :7) ;如果入侵者动作很 轻(附肢没有变形或毁坏) ,甲虫则逃跑[22 ] 。并 且发现经过 1 次蜕皮后其相应部位能够再生。 114 自残部位 凡是能够肢体自残的种类均有特定的自残 部位 ,一般位于与身体连接最少的部分。如以 足自残的昆虫自残部位为转节与股节之间 ,以 尾须和鳃自残的昆虫自残部位为尾须和鳃的基 部 ,贝氏安角蝉 Anchistrotus besckii ( Germar) 的自 残部位在前胸背板膨大部分的基部 ,而棕拟花 蚤 Scraptia fuscula (Müller) 的幼虫腹末附器自残 部位在第 8 腹节和附器基部之间。在身体主体 和被残器官间有一层膜能有效阻止自残后血液 外流。 2 引起自残的原因 引起昆虫自残的原因主要有以下几个方 面。 211 捕食者和人类的威胁 这是昆虫自残最 主要的原因。当自残者被人类或捕食者握住或 噙住自残器官或当其受到捕食者威胁时 ,有自 残能力的昆虫会很快断掉自残器官而逃跑。如 果您捉到蝗科或螽斯科等直翅目昆虫 ,用食指 和拇指握住其一只或一对后足 ,被握的足很快 会被自残者断掉。 212 意外伤害 当蟋蟀等直翅目昆虫的后足 的一部分受到伤害 (如胫节断掉) 时 ,这些直翅 目昆虫常常会从转节和股节的连接处把大部分 后足断掉。 213 接触毒物或杀虫剂 首次用化学成分诱 导昆虫肢体自残的报道见于 1943 年 , Emmel 用 DDT处理花翅按蚊 Anopheles maculipennis Meigen 能使按蚊后足自残[8 ] 。1944 年 , Wiesmann 和 Fenjves 报道了尖音库蚊 Culex pipiens Linnaeus、 麦蛾 Sitotroga cerealella (Olivier) 及 一 种 尺 蠖 Cheimatobia brumata (Linnaeus) 接触 DDT 后成虫 后足自残的现象[9 ] 。Brousse2Gaury 发现用氰化 钾注入或用甲醛熏蒸均能引起家蟋蟀的后足自 残 [7 ] 。Uvarov 也曾提及当把蝗虫放入毒瓶时 , 蝗虫也会发生后足自残[4 ] 。20 世纪 80 年代以 来 ,Moore 及其合作者对小菜蛾 Plutella xylostella (Linnaeus) [10 ,11 ] 、Ortego 和 Bowers 对美洲蚱蜢 Schistocerca americana (Drury) 的药剂诱导自残做 了比较详细的研究[23 ] 。 214 种群密度过大 对一些具有自相残杀习 性的蜻蜓目稚虫而言 ,在密度较高时尾鳃增加 了被同类捕食的机会 ,因此在实验条件下随着 稚虫密度的增大 , 尾鳃自残的比例也就上 升 [15 ,16 ] 。 此外 ,在实验条件下 ,一些物理刺激 (如机 械压力、温度变化和电击等) 也能引起蝗虫和蟋 · 282 · 昆虫知识 ENTOMOLOGICAL KNOWLEDGE 2003 40(3) © 1994-2007 China Academic Journal Electronic Publishing House. All rights reserved. http://www.cnki.net

2003 40(3)昆虫知识ENTOMOLOGICALKNOWLEDGE·283:蜂的后足自残(4,7)]。捕食者残杀的数量比全尾鳃的未龄稚虫多2倍。相对于有尾鳃的稚虫来说,无尾鳃的稚虫3肢体自残对昆虫的影响在受到威胁时不易逃离,逃离的比例前者明显3.1对昆虫的有利影响肢体自残的实质是大于后者(前者为29%,后者仅为9%)。在这昆虫以栖牲局部利益换取个体生存的机会,这一例子中,由自残造成的长期不利几乎与短期利益抵消15]。对个体生命的延续和种群的繁荣有着积极的意义。Paul认为自残的主要好处在于利用这个机昆虫的自残挽救了其生命。但是,就已知会身体的其他部分可以逃离是非之地。面临捕的自残的昆虫而言尚未发现某一类昆虫所有食者或不利条件,自残能使自残者受害的危险实验个体都发生自残,也就是说只有一定比例降到最低。角蝉被鸟类叼住膨大的前胸背板的个体发生自残。时,其膨大部分可能被捕食者吃掉,但身体的其4自残与再生的关系他部分可趁机逃生。对于小菜蛾而言,它的保护机制同样也是如此。小菜蛾接触农药后的反昆虫幼虫期发生肢体自残,有些种类经过映过程为:昏一自残死亡(或苏醒)。如果蜕皮其相应部分还能够再生(regeneration)[6),如让小菜蛾在氰戊菊酯残留上(250ng/cm)爬行蜻蜓目稚虫的尾腮自残后能再生,但再生肢比1min,48h后48%的小菜蛾断掉1个或2个后原肢短小:昆虫成虫期发生自残则再生不能进足发生自残的小菜蛾死亡率18%,没有发生行,如小菜蛾节接触农药残留后足的自残,再自残的小菜蛾死亡率31%。3h后,发生断肢生无法进行。昆虫的断肢再生是一个非常复杂的小菜蛾体内的农药残留量及农药代谢物浓度的生命现象,涉及到形态学、生理学、分子生物比没发生断肢的小菜蛾的相应的量低21%。学、发育遗传学等问题,我们将另文讨论经测定,残掉的小菜蛾后足中农药的浓度及代谢物的量比自残后的体内相应数量高10倍。致谢承蒙西北农林科技大学袁锋教授、捷克它以自残的方式排出体内的农药剂量,以避免共和国科学院昆虫研究所P.Svacha博士及日农药影响身体的其它部分。后足自残后,小菜本国立科学博物馆友国雅章博士惠赠资料。中蛾利用剩余足行走没有困难,照样能够飞行、交国农业大学昆虫学系王心丽副教授、刘志琦副配和产生可育后代""。教授等讨论有关问题,谨此铭谢。3.2自残给剩余肢体带来不利影响自残可参考文献以使昆虫逃避捕食者和其他不利条件也为以1Fredericq L, Archs. Zool. Exp. Gen. ,I :413~426.后的生活带来一系列负面影响。蜻蜓目稚虫虫2Wood F.D.,Wood HE.J.Exp.Zool.,1932,62 :1~155.口密度大时有自相残杀的特性,无尾鳃末龄稚3MeVean A. Comp. Biochem. Phmsiol. ,1974,51A :497~505虫常常“逃得了一时逃不了一世”。对于蜻蜓目4Uvarov B. P. Grasshoppers and Locusts. A Handbook of General稚虫而言,尾鳃的主要功能在于气体交换,此虫Acridology,Vol. 2. Cambridge:Cambridge University Press1977.385,并不适合于在氧气浓度低的条件下生存,有尾5Steiner A.L. Pyche ,1968 ,75 :256~263鳃的稚虫和无尾鳃的稚虫个体在水中的纵向分6Bulliere D. ,Bulliere F. In: Kerkut G A. ,Glbert L. I, (eds.)Comprehensive Insect Physiology ,Biochemistry and Pharmacolo布很不相同,无尾鳃的个体趋向于更高浓度的gy,Vol. 2. England : Pergamon Press,1985.371~424氧气的水流[15,16]。同时尾鳃位于腹部末端的游7Brousse-Caury M. P. Bull. Biol, Fr. Belg- ,1958 ,92 55~63动部位,它的存在保证了稚虫游动的能力。无8Emmel L.Z.Hyg.Zool.,1943,35 :119~124.oWiesmann R. ,Fenjves P. Mitt. Scinreiz. Entomol. Ges.,1944,尾腮时稚虫似蛇状婉蜓游动,并且效率低,这种19:179~184低效率对躲避捕食者是不利的。在实验过程中101Mbore A, , Tabashnik B. E. J. Econ. Entomol. ,1989 ,82 (2) :发现:尾鳃少的末龄稚虫被同种个体或它种的381~384.D1994-2007ChinaAcademicJournalElectronicPublishingHouse.All rightsreserred.htp:/hn.cnki.nen
蟀的后足自残[4 ,7 ] 。 3 肢体自残对昆虫的影响 311 对昆虫的有利影响 肢体自残的实质是 昆虫以牺牲局部利益换取个体生存的机会 ,这 对个体生命的延续和种群的繁荣有着积极的意 义。Paul 认为自残的主要好处在于利用这个机 会身体的其他部分可以逃离是非之地。面临捕 食者或不利条件 ,自残能使自残者受害的危险 降到最低。角蝉被鸟类叼住膨大的前胸背板 时 ,其膨大部分可能被捕食者吃掉 ,但身体的其 他部分可趁机逃生。对于小菜蛾而言 ,它的保 护机制同样也是如此。小菜蛾接触农药后的反 映过程为 :昏厥 —自残 —死亡 (或苏醒) 。如果 让小菜蛾在氰戊菊酯残留上 (250 ngΠcm 2 ) 爬行 1 min ,48 h 后 48 %的小菜蛾断掉 1 个或 2 个后 足 ;发生自残的小菜蛾死亡率 18 % ,没有发生 自残的小菜蛾死亡率 31 %。3 h 后 ,发生断肢 的小菜蛾体内的农药残留量及农药代谢物浓度 比没发生断肢的小菜蛾的相应的量低 21 %。 经测定 ,残掉的小菜蛾后足中农药的浓度及代 谢物的量比自残后的体内相应数量高 10 倍。 它以自残的方式排出体内的农药剂量 ,以避免 农药影响身体的其它部分。后足自残后 ,小菜 蛾利用剩余足行走没有困难 ,照样能够飞行、交 配和产生可育后代[10 ,11 ] 。 312 自残给剩余肢体带来不利影响 自残可 以使昆虫逃避捕食者和其他不利条件 ,也为以 后的生活带来一系列负面影响。蜻蜓目稚虫虫 口密度大时有自相残杀的特性 ,无尾鳃末龄稚 虫常常“逃得了一时逃不了一世”。对于蜻蜓目 稚虫而言 ,尾鳃的主要功能在于气体交换 ,此虫 并不适合于在氧气浓度低的条件下生存 ,有尾 鳃的稚虫和无尾鳃的稚虫个体在水中的纵向分 布很不相同 ,无尾鳃的个体趋向于更高浓度的 氧气的水流[15 ,16 ] 。同时尾鳃位于腹部末端的游 动部位 ,它的存在保证了稚虫游动的能力。无 尾腮时稚虫似蛇状蜿蜒游动 ,并且效率低 ,这种 低效率对躲避捕食者是不利的。在实验过程中 发现 :尾鳃少的末龄稚虫被同种个体或它种的 捕食者残杀的数量比全尾鳃的末龄稚虫多 2 倍。相对于有尾鳃的稚虫来说 ,无尾鳃的稚虫 在受到威胁时不易逃离 ,逃离的比例前者明显 大于后者 (前者为 29 % ,后者仅为 9 %) 。在这 一例子中 ,由自残造成的长期不利几乎与短期 利益抵消[15 ] 。 昆虫的自残挽救了其生命。但是 ,就已知 的自残的昆虫而言 ,尚未发现某一类昆虫所有 实验个体都发生自残 ,也就是说只有一定比例 的个体发生自残。 4 自残与再生的关系 昆虫幼虫期发生肢体自残 ,有些种类经过 蜕皮其相应部分还能够再生(regeneration) [6 ] ,如 蜻蜓目稚虫的尾腮自残后能再生 ,但再生肢比 原肢短小 ;昆虫成虫期发生自残则再生不能进 行 ,如小菜蛾跗节接触农药残留后足的自残 ,再 生无法进行。昆虫的断肢再生是一个非常复杂 的生命现象 ,涉及到形态学、生理学、分子生物 学、发育遗传学等问题 ,我们将另文讨论。 致谢 承蒙西北农林科技大学袁锋教授、捷克 共和国科学院昆虫研究所 P. Svácha 博士及日 本国立科学博物馆友国雅章博士惠赠资料。中 国农业大学昆虫学系王心丽副教授、刘志琦副 教授等讨论有关问题 ,谨此铭谢。 参 考 文 献 1 Frédéricq L. Archs. Zool. Exp. Gen. ,1 :413~426. 2 Wood F. D. , Wood H. E. J . Exp. Zool. , 1932 ,62 : 1~ 155. 3 McVean A. Comp. Biochem. Physiol. ,1974 ,51A :497~505. 4 Uvarov B. P. Grasshoppers and Locusts. A Handbook of General Acridology , Vol. 2. Cambridge : Cambridge University Press , 1977. 385. 5 Steiner A.L. Psyche ,1968 ,75 :256~263. 6 Bulliére D. ,Bulliére F. In : Kerkut G. A. ,Gilbert L. I. (eds. ) . Comprehensive Insect Physiology ,Biochemistry and Pharmacolo2 gy ,Vol. 2. England : Pergamon Press ,1985. 371~424. 7 Brousse2Gaury M. P. Bull. Biol. Fr. Belg. ,1958 ,92 :55~63. 8 Emmel L. Z. Hyg. Zool. ,1943 ,35 :119~124. 9 Wiesmann R. ,Fenjves P. Mitt. Schweiz. Entomol. Ges. ,1944 , 19 :179~184. 10 Moore A. , Tabashnik B. E. J . Econ. Entomol. ,1989 ,82 (2) : 381~384. 2003 40(3) 昆虫知识 ENTOMOLOGICAL KNOWLEDGE · 382 · © 1994-2007 China Academic Journal Electronic Publishing House. All rights reserved. http://www.cnki.net

.284.昆虫知识ENTOMOLOGICALKNOWLEDGE2003.40(3)11More A., Tabashnik B. E., Stark J. D. J. Econ. Emomo.21 Carlberg U. Zool. Are. ,1992 ,228(5/6) : 229~237.198982(5):1295~129822Svocha P. In : Pakaluk J. ,Slipinski S. A. (eds. ) ,Biology ,Phy12Strlimpel H. Ent. Mit. Zod. Mis. Hambug,1983,7(119) :391logeny,and Classification of Coleoptera. Warszawa : Muzeum I I~396.stytut Zoologi PAN,1995.473~48913Rietschel S. Entomol. Gener. ,1987,12(4) :209~22023Ortego F.,Bowers W.S. Experientia ,1996,52:42~5014Tabashnik B. E.,Cushing N.L. Johnson M. W. J. Eoon. Ento24.Matthews R. W.,Matthews J. R. Insect Behavior. New York :John Wiley & Son ,1978.507.mol.,1987,80:1 091~1 099.15Robinson J. V., Hayworth D. A., Harvey M. V: Am. Midl.25Wigglesworth V. B. The Principles of Insect Physiology. 7th edChapman and Hall,1972. 827.Nat.,1991,125240~24416RobinsonJ.V.,Shaffer L, R. Oecologia,1991,87 :1~7.26Wood T.K.,Mbrris G K Can.Entomol.,1974,106:143~17 Stoks R. Beh. Ecol. Sociobiol. ,1999 ,47:70~75.148.1827Wood T. K Proc. Entomol. Soc. Wash. .1975,77 :78~82StoksR.J.Zool.,1999,247:269~2731928Stoks R.,De Block M. , Van Cossum H. , Valck F, ,LaumersWoodT.KAm.RevEntamol.,1993,38:409~435.K.,Oecologia,1999,120:87~9129田润刚,袁锋.西北农业大学学报,1995,26(3):23~2720Carlberg U. Zool. Arz. ,1986 ,217(1/2) : 39~53.30袁锋.昆虫分类学报,1998,20(1):1~17.昆虫的睡眠与生物钟元文琪(瑞典隆德大学生态学系)Sleep and biological clock in insects Wenqi Rosen(Department gf Ecology, Lund University,Lund ,Sweden)AbstracetSeveral evidences have indicated that rest in insects is a sleeplike state. The present article summarizesthose findings which demonstrated how insect rest behaviour met the criteria for sleep ,and the similarities in the phystological mechanisms of sleep between mammals and insects. In addition,the paper also elucidates the importance of brological clock genes in the regulationof insect seep.Key wordsinsect sleep, biological clock摘要近年来,越来越多的证据表明昆虫也要睡眠。该文介绍这方面的研究发现,包括确定昆虫睡眠的标准以及昆虫与哺乳动物的睡眠在生理机制上的相似性,最后阐述了生物钟基因对昆虫睡眠的调节作用。关键词昆虫的睡眠,生物钟众所周知,人体需要睡眠,缺乏了睡眠,体一个调节体内睡眠平衡机制即当动物睡眠被剥内的免疫系统、内分泌功能都会紊乱,大脑就不夺了几个小时后,它以后的睡眠时间明显地增长!。借助于录像机,科学家们对一系列昆虫如能正常地思维和工作,长期缺乏睡眠严重的甚至会导致死亡。很久以来,睡眠一直被认为只蟑螂、蟋蜂、果蝇和蜜蜂每天24h的活动进行了是人类及高级哺乳动物和一些鸟类专有的特长期观察,他们发现这些昆虫的“起居饮食”其实征。但近几年来,越来越多的证据表明那些低很有规律,它们每天在一个固定的时间到一个固级的无脊椎动物,如昆虫也要睡觉。定的地方呆着不动,长达几个小时。在这期间,确定一个动物是否有睡眠行为并不只看它每种昆虫还有专门的姿势,如蜜蜂,它们的触角是不是不动、对外界刺激有无反应。科学家们有在休息时总是下垂着,而在其它时间,触角总是以下几个公认的标准:(1)在一定的时间内长期竖着不停地上下摆动(图12)(2]。地不动:(2)有一个特定的姿势和专门的场所:收稿日期:2002-11-28,接收日期:2003-01-27(3)在不动时对外界的刺激反应阈值增加:(4)有D1994-2007ChinaAcademicJournalElectronicPublishingHouse.AllrightsreservedLhtp:/hwww.cnki.ner
11 Moore A. , Tabashnik B. E. , Stark J . D. J . Econ. Entomo. , 1989 ,82(5) : 1 295~1 298. 12 Strümpel H. Ent. Mitt. Zool. Mus. Hamburg ,1983 ,7 (119) :391 ~396. 13 Rietschel S. Entomol. Gener. ,1987 ,12(4) :209~220. 14 Tabashnik B. E. ,Cushing N. L. ,Johnson M. W. J . Econ. Ento2 mol. ,1987 ,80 :1 091~1 099. 15 Robinson J . V. , Hayworth D. A. , Harvey M. V. Am. Midl. Nat. ,1991 ,125 :240~244. 16 Robinson J . V. ,Shaffer L. R. Oecologia ,1991 ,87 :1~7. 17 Stoks R. Beh. Ecol. Sociobiol. ,1999 ,47 :70~75. 18 Stoks R. J . Zool. ,1999 ,247 :269~273. 19 Stoks R. ,De Block M. , Van Gossum H. , Valck F. ,Lauwers K. , Oecologia ,1999 ,120 :87~91. 20 Carlberg U. Zool. Anz. ,1986 ,217(1Π2) : 39~53. 21 Carlberg U. Zool. Anz. ,1992 ,228(5Π6) : 229~237. 22 Svácha P. In : Pakaluk J . ,Slipinski S. A. (eds. ) ,Biology , Phy2 logeny ,and Classification of Coleoptera. Warszawa : Muzeum I In2 stytut Zoologii PAN ,1995. 473~489. 23 Ortego F. ,Bowers W. S. Experientia ,1996 ,52 :42~50. 24 Matthews R. W. , Matthews J . R. Insect Behavior. New York : John Wiley & Son ,1978. 507. 25 Wigglesworth V. B. The Principles of Insect Physiology. 7th ed. Chapman and Hall ,1972. 827. 26 Wood T. K. ,Morris G. K. Can. Entomol. ,1974 ,106 : 143~ 148. 27 Wood T. K. Proc. Entomol. Soc. Wash. ,1975 ,77 :78~82. 28 Wood T. K. Ann. Rev. Entomol. ,1993 ,38 :409~435. 29 田润刚 ,袁锋. 西北农业大学学报 ,1995 ,26 (3) :23~27. 30 袁锋. 昆虫分类学报 ,1998 ,20 (1) :1~17. 昆虫的睡眠与生物钟 兀文琪 (瑞典隆德大学生态学系) Sleep and biological clock in insects. Wenqi Rosen( Department of Ecology , Lund University ,Lund ,Sweden) . Abstract Several evidences have indicated that rest in insects is a sleep2like state. The present article summarizes those findings which demonstrated how insect rest behaviour met the criteria for sleep ,and the similarities in the physi2 ological mechanisms of sleep between mammals and insects. In addition ,the paper also elucidates the importance of bi2 ological clock genes in the regulation of insect sleep. Key words insect sleep , biological clock 摘 要 近年来 ,越来越多的证据表明昆虫也要睡眠。该文介绍这方面的研究发现 ,包括确定昆虫睡眠 的标准以及昆虫与哺乳动物的睡眠在生理机制上的相似性 ,最后阐述了生物钟基因对昆虫睡眠的调节 作用。 关键词 昆虫的睡眠 , 生物钟 收稿日期 :2002211228 ,接收日期 :2003201227 众所周知 ,人体需要睡眠 ,缺乏了睡眠 ,体 内的免疫系统、内分泌功能都会紊乱 ,大脑就不 能正常地思维和工作 ,长期缺乏睡眠 ,严重的甚 至会导致死亡。很久以来 ,睡眠一直被认为只 是人类及高级哺乳动物和一些鸟类专有的特 征。但近几年来 ,越来越多的证据表明那些低 级的无脊椎动物 ,如昆虫也要睡觉。 确定一个动物是否有睡眠行为并不只看它 是不是不动、对外界刺激有无反应。科学家们有 以下几个公认的标准: (1) 在一定的时间内长期 地不动; (2) 有一个特定的姿势和专门的场所; (3)在不动时对外界的刺激反应阈值增加; (4) 有 一个调节体内睡眠平衡机制 ,即当动物睡眠被剥 夺了几个小时后 ,它以后的睡眠时间明显地增 长 [1] 。借助于录像机 ,科学家们对一系列昆虫如 蟑螂、蟋蟀、果蝇和蜜蜂每天 24 h 的活动进行了 长期观察 ,他们发现这些昆虫的“起居饮食”其实 很有规律 ,它们每天在一个固定的时间到一个固 定的地方呆着不动 ,长达几个小时。在这期间 , 每种昆虫还有专门的姿势 ,如蜜蜂 ,它们的触角 在休息时总是下垂着 ,而在其它时间 ,触角总是 竖着不停地上下摆动(图 1 ,2) [2] 。 · 482 · 昆虫知识 ENTOMOLOGICAL KNOWLEDGE 2003 40(3) © 1994-2007 China Academic Journal Electronic Publishing House. All rights reserved. http://www.cnki.net