
第七章脂类代谢 脂类化合物包括甘油三酯和类脂质。甘油三酯是生物体的主要储能物质,类脂质大都 是细胞的重要结构物质和生理活性物质。本章主要介绍脂类化合物的概念、类别、分布和 生理功能,并重点讨论中性脂肪 三酰甘油在有机体内的合成和降解代谢。 脂类(lipids)是指一类在化学组成和结构上有很大差异,但都有一个共同特性,即不 溶于水而易溶于乙醚、氯仿等非极性溶剂的物质。通常脂类可按不同组成分为五类,即单 纯脂、复合 ,萜类和类固醇及其衍生物 ,衍生脂类及结合脂类 脂类物质具有重婴的生物功能。脂肪是生物体的能量提供者。1g脂肪彻底氧化可放出 46.5 kJ/mol能量,比1g糖或蛋白质放出的能量大一倍以上,因此脂肪是生物体内贮藏能量 最多的物质。 每1g蛋白质、糖和脂肪所产生的代谢能量比较为: 1g蛋白质 20.5km0 1g糖 20.5kJ/mo 1g脂肪 46.5kJ/mol 这样大的能量差异是因为脂肪是非极性的,它以近于无水的形式储藏者,而糖类和蛋 白质均具有极性,它们以高度水合形式储藏者。1g干燥的糖元约结合2g水,所以实际上 ,这就说明了为什么在进化过程中选择 脂肪也是组成生物体的重要成分,如磷脂是构成生物膜的重要组分,油脂是机体代谢 所需燃料的贮存和运输形式。脂类物质也可为动物机体提供溶解于其中的必需脂肪酸和脂 溶性维生素。某些茜类及类固醇类物质如维生素A、D、E、K、胆酸及固醇类激素具有营 养、代谢及调节功能。有机体表面的脂类物质有防止机械损伤与防止热量散发等保护作用。 脂类作为细 的表面物质 与细胞识别和种特异性以及组织免疫等有密切关系 脂类代谢主要讨论脂类在有机体内的降解和合成过程。了解脂类代谢对农业、工业 医学等方面都有重要的意义。例如种子的发芽率直接和种子的脂类代谢有关:又如利用微 生物氧化石油中脂肪烃、工业生产低凝点油及其他化工产品:脂蛋白异常和威胁人类健康 的冠心病等都与脂肪代谢关系密切。 第一节脂肪的降解 一、脂肪的酶促降解 脂肪即脂肪酸的甘油三酯(triacyl glycerol)是脂类中含量最丰富的一大类,它是甘油 的三个羟基和三个脂肪酸分子缩合、失水后形成的酯,是植物和动物细胞贮脂的主要组分
225 第七章 脂类代谢 脂类化合物包括甘油三酯和类脂质。甘油三酯是生物体的主要储能物质,类脂质大都 是细胞的重要结构物质和生理活性物质。本章主要介绍脂类化合物的概念、类别、分布和 生理功能,并重点讨论中性脂肪 三酰甘油在有机体内的合成和降解代谢。 脂类(lipids)是指一类在化学组成和结构上有很大差异,但都有一个共同特性,即不 溶于水而易溶于乙醚、氯仿等非极性溶剂的物质。通常脂类可按不同组成分为五类,即单 纯脂、复合脂、萜类和类固醇及其衍生物、衍生脂类及结合脂类。 脂类物质具有重要的生物功能。脂肪是生物体的能量提供者。1g 脂肪彻底氧化可放出 46.5kJ/mol 能量,比 1g 糖或蛋白质放出的能量大一倍以上,因此脂肪是生物体内贮藏能量 最多的物质。 每 1g 蛋白质、糖和脂肪所产生的代谢能量比较为: 1g 蛋白质 20.5kJ/mol 1g 糖 20.5kJ/mol 1g 脂肪 46.5kJ/mol 这样大的能量差异是因为脂肪是非极性的,它以近于无水的形式储藏着,而糖类和蛋 白质均具有极性,它们以高度水合形式储藏着。1g 干燥的糖元约结合 2g 水,所以实际上 1g 脂肪所贮存的能量为 1g 水合糖元贮存的六倍多。这就说明了为什么在进化过程中选择 脂肪作为能量的储备形式。 脂肪也是组成生物体的重要成分,如磷脂是构成生物膜的重要组分,油脂是机体代谢 所需燃料的贮存和运输形式。脂类物质也可为动物机体提供溶解于其中的必需脂肪酸和脂 溶性维生素。某些萜类及类固醇类物质如维生素 A、D、E、K、胆酸及固醇类激素具有营 养、代谢及调节功能。有机体表面的脂类物质有防止机械损伤与防止热量散发等保护作用。 脂类作为细胞的表面物质,与细胞识别和种特异性以及组织免疫等有密切关系。 脂类代谢主要讨论脂类在有机体内的降解和合成过程。了解脂类代谢对农业、工业、 医学等方面都有重要的意义。例如种子的发芽率直接和种子的脂类代谢有关;又如利用微 生物氧化石油中脂肪烃、工业生产低凝点油及其他化工产品;脂蛋白异常和威胁人类健康 的冠心病等都与脂肪代谢关系密切。 第一节 脂肪的降解 一、脂肪的酶促降解 脂肪即脂肪酸的甘油三酯(triacyl glycerol)是脂类中含量最丰富的一大类,它是甘油 的三个羟基和三个脂肪酸分子缩合、失水后形成的酯,是植物和动物细胞贮脂的主要组分

脂肪降解的第一步是水解成甘油和脂肪酸,此反应由脂肪酶(简称脂酶,las)催化。组 织中有三种脂肪酶:即脂肪酶、甘油二酯脂肪酶和甘油单酯脂肪酶,逐步把甘油三酯水解 成甘油和脂肪酸。这三种酶水解步骤为: o CH:-0-C-R 十品0脂防晓 0H4C-0-C-R B- H CH:O-C-R H:COH 甘油三 脂肪酸 甘油二面 甘油二酯脂防酶 甘油单酯脂肪酶 H:C-OH -0 H- R.COOH R COOH H.COH HC-OH 脂肪酸 脂肪酸 甘油单酯 甘油 在人和动物消化道内有脂肪醇,分解食物中的脂肪:以后甘油和脂肪酸在组织内再进 一步氧化分解。植物也有类似的脂肪消化作用。如油料作物的种子萌发时,种子内脂肪酶 活力增加,促使脂肪发生分解。凡能利用脂肪的微生物也都有脂肪酶,生产春雷霉素的培 养基中需含有一定配出的物油,说明春需露黄能铭产生脂肪酶,所以能利用植物油,假 丝酵母、圆酵母等都能产生较多的脂肪酶, 已经利用它们作为制造脂肪酶制剂的原 二、甘油的降解与转化 甘油先与ATP作用,在甘油激酶(glycerol kinase)催化下生成a-磷酸甘油。然后再 被氧化生成磷酸二羟丙酮,再经异构化,生成3-磷酸甘油醛,然后可经糖酵解途径转化成 丙酮酸,进入三羧酸循环而彻底氧化,或经过糖异生途径合成糖元。因此甘油代谢和糖代 谢的关系极为密切。甘油转化成磷酸二羟丙酮以及与糖的相互转变关系如下: 226
226 脂肪降解的第一步是水解成甘油和脂肪酸,此反应由脂肪酶(简称脂酶,lipase)催化。组 织中有三种脂肪酶:即脂肪酶、甘油二酯脂肪酶和甘油单酯脂肪酶,逐步把甘油三酯水解 成甘油和脂肪酸。这三种酶水解步骤为: 在人和动物消化道内有脂肪酶,分解食物中的脂肪;以后甘油和脂肪酸在组织内再进 一步氧化分解。植物也有类似的脂肪消化作用。如油料作物的种子萌发时,种子内脂肪酶 活力增加,促使脂肪发生分解。凡能利用脂肪的微生物也都有脂肪酶,生产春雷霉素的培 养基中需含有一定配比的植物油,说明春雷霉菌能够产生脂肪酶,所以能利用植物油,假 丝酵母、圆酵母等都能产生较多的脂肪酶,工业上已经利用它们作为制造脂肪酶制剂的原 料。 二、甘油的降解与转化 甘油先与 ATP 作用,在甘油激酶(glycerol kinase)催化下生成α-磷酸甘油。然后再 被氧化生成磷酸二羟丙酮,再经异构化,生成 3-磷酸甘油醛,然后可经糖酵解途径转化成 丙酮酸,进入三羧酸循环而彻底氧化,或经过糖异生途径合成糖元。因此甘油代谢和糖代 谢的关系极为密切。甘油转化成磷酸二羟丙酮以及与糖的相互转变关系如下:

CH.OH ATP ADP CH.OH NAD NADH+H CHOH 口-醉酸甘袖脱氢酶 一葡萄培一一一一一糖元 CH2OH c=0 CHOP 两酸二羟丙扇 CO:+H.O+ATP 三、脂肪酸的氧化分解 (一)饱和脂肪酸的B-氧化作用 1.B氧化作用的概念 脂肪酸的B氧化作用是指脂肪酸在一系列酶的作用下,在ā,B碳原子之间断裂,B 碳原子氧化成羧基,生成含2个碳原子的乙酰-C0A和较原来少2个碳原子的脂肪酸。脂 肪酸的B氧化过程是在线粒体中进行的 B氧化作用最初是根据动物实验提出来的一个学说。通过制备一系列的苯(基) 脂(肪)酸,即脂肪酸的甲基(-CH)上的一个氢原子被苯基取代而成的苯脂酸,再将它 们饲喂动物。在动物体内,苯基不被破坏,而是通过解毒机制,形成无毒性的衍生物,从 尿中排出。鉴定尿中含苯基的化合物,可以推测脂肪酸在体内的分解途径。将带有苯基的 双数和单数碳的脂肪酸喂给狗吃,从尿中分离到两种含苯基的化合 种是由苯甲酸和 甘氨酸缩合而成的马尿酸,另一种是由苯乙酸和甘氨酸缩合而成的苯乙尿酸。凡是吃了 数碳苯脂酸的狗,尿中的苯基化合物为苯乙尿酸,凡是吃了单数碳苯脂酸的狗,尿中的苯 基化合物为马尿酸。由于每次断下1个碳或断下3个碳都不符合实验结果,Koop认为脂 肪酸在体内氧化时每次都断下1个二碳物(见图7-1)。他在1904年提出的B-氧化学说, 至今仍旧是正确的」 氧化作用并不是一步完成的,而是要经过活化、转运,然后再进入氧化过程。 2.脂肪酸的活化 脂肪酸在进行B-氧化降解前,在细胞质内必须先被激活成脂酰-CA,该反应由脂酰 -CoA合成酶(acyl-CoA synthetase)催化,需要ATP和CoA参与,总反应为: 27
227 三、脂肪酸的氧化分解 (一)饱和脂肪酸的β-氧化作用 1. β-氧化作用的概念 脂肪酸的β-氧化作用是指脂肪酸在一系列酶的作用下,在α,β-碳原子之间断裂,β -碳原子氧化成羧基,生成含 2 个碳原子的乙酰-CoA 和较原来少 2 个碳原子的脂肪酸。脂 肪酸的β-氧化过程是在线粒体中进行的。 β-氧化作用最初是根据动物实验提出来的一个学说。通过制备一系列的ω-苯(基) 脂(肪)酸,即脂肪酸的甲基(-CH3)上的一个氢原子被苯基取代而成的苯脂酸,再将它 们饲喂动物。在动物体内,苯基不被破坏,而是通过解毒机制,形成无毒性的衍生物,从 尿中排出。鉴定尿中含苯基的化合物,可以推测脂肪酸在体内的分解途径。将带有苯基的 双数和单数碳的脂肪酸喂给狗吃,从尿中分离到两种含苯基的化合物。一种是由苯甲酸和 甘氨酸缩合而成的马尿酸,另一种是由苯乙酸和甘氨酸缩合而成的苯乙尿酸。凡是吃了双 数碳苯脂酸的狗,尿中的苯基化合物为苯乙尿酸,凡是吃了单数碳苯脂酸的狗,尿中的苯 基化合物为马尿酸。由于每次断下 1 个碳或断下 3 个碳都不符合实验结果,Knoop 认为脂 肪酸在体内氧化时每次都断下 1 个二碳物(见图 7-1)。他在 1904 年提出的β-氧化学说, 至今仍旧是正确的。 β-氧化作用并不是一步完成的,而是要经过活化、转运,然后再进入氧化过程。 2. 脂肪酸的活化 脂肪酸在进行β-氧化降解前,在细胞质内必须先被激活成脂酰-CoA,该反应由脂酰 -CoA 合成酶(acyl-CoA synthetase)催化,需要 ATP 和 CoA 参与,总反应为:
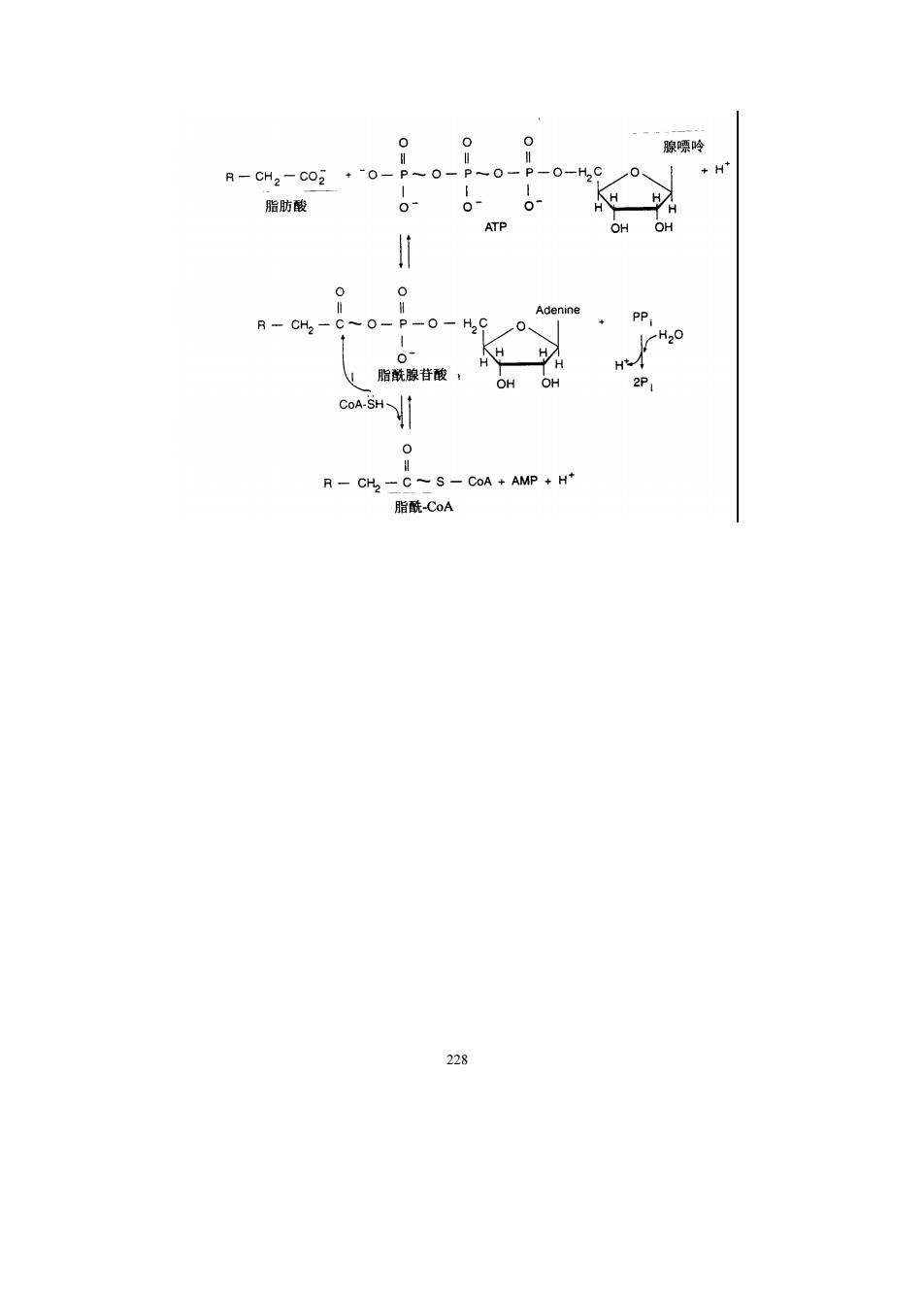
吟 R-CH2-C02 +0- 脂肪酸 CH,- C -S-CoA AMP +H* 脂酰CoA 228
228
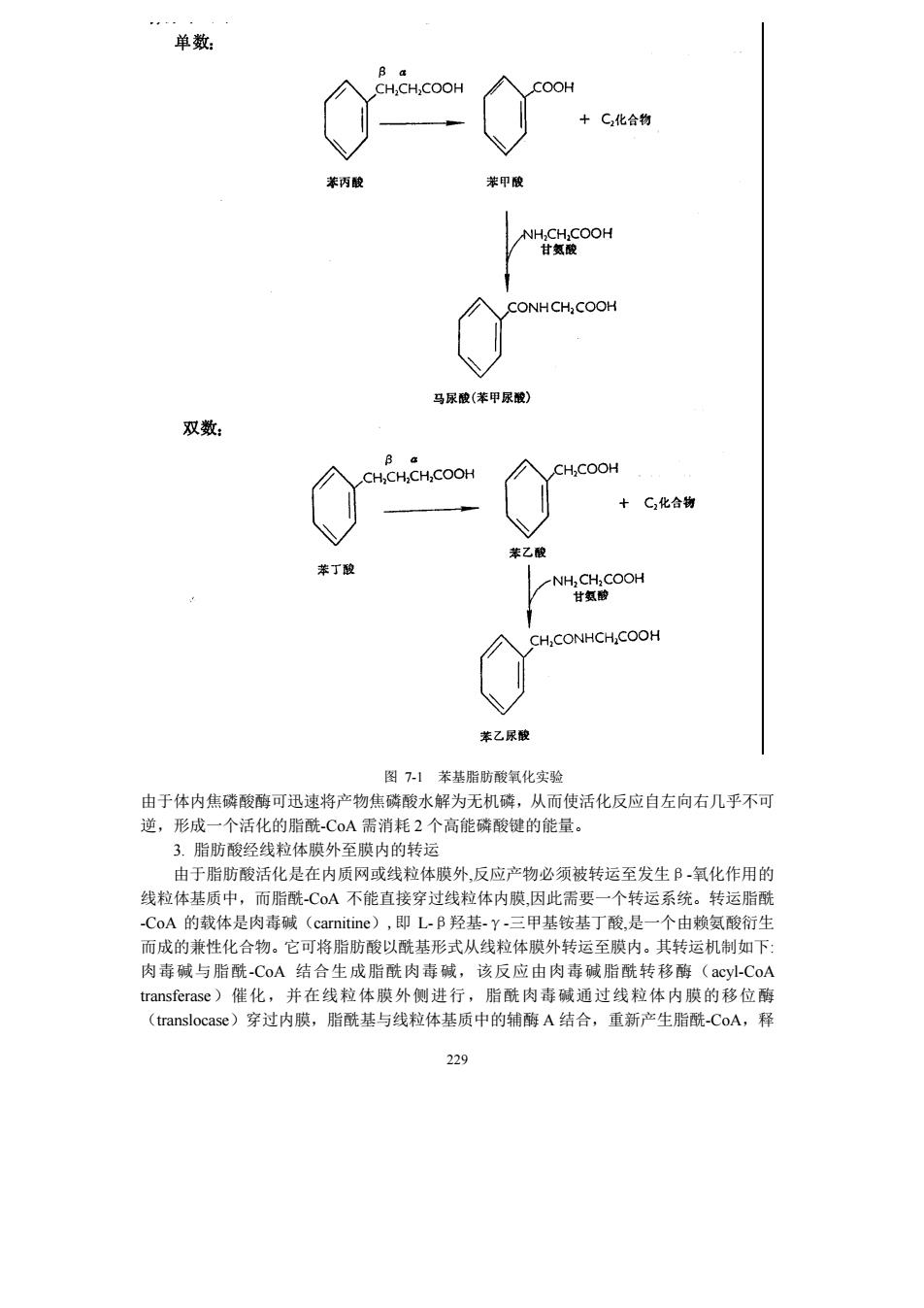
单数 CH.CH,COOH C,化合物 CONHCH.COOH 马尿酸(甲尿酸) 双数 a CH.CH,CH,COOH C化合物 NHCOOH CH,CONHCH.COOH 苯乙尿酸 图7-1苯基脂防酸氧化实验 由于体内焦磷酸酶可迅速将产物焦磷酸水解为无机磷,从而使活化反应自左向右几乎不可 逆,形成一个活化的脂酰C0A需消耗2个高能磁酸辩的能量。 3。脂防酸经线粒体腻外至膜内的转运 由于脂肪酸活化是在内质网或线粒体膜外,反应产物必须被转运至发生B氧化作用的 线粒体基质中,而脂酰-C0A不能直接穿过线粒体内膜,因此需嬰一个转运系统。转运脂酰 -CoA的载体是肉毒碱(carnitine),即L-B羟基-Y-三甲基铵基丁酸,是一个由赖氨酸衍生 而成的兼性化合物。它可将脂肪酸以酰基形式从线粒体膜外转运至膜内。其转运机制如下 肉毒碱与脂酰-CA结合生成脂酰肉毒碱,该反应由肉毒碱脂酰转移酶 transferase)催化,并在线粒体膜外侧进行,脂酰肉毒碱通过线粒体内膜的移位酶 (translocase)穿过内膜,脂酰基与线粒体基质中的辅醇A结合,重新产生脂酰-CoA,释 229
229 图 7-1 苯基脂肪酸氧化实验 由于体内焦磷酸酶可迅速将产物焦磷酸水解为无机磷,从而使活化反应自左向右几乎不可 逆,形成一个活化的脂酰-CoA 需消耗 2 个高能磷酸键的能量。 3. 脂肪酸经线粒体膜外至膜内的转运 由于脂肪酸活化是在内质网或线粒体膜外,反应产物必须被转运至发生β-氧化作用的 线粒体基质中,而脂酰-CoA 不能直接穿过线粒体内膜,因此需要一个转运系统。转运脂酰 -CoA 的载体是肉毒碱(carnitine),即 L-β羟基-γ-三甲基铵基丁酸,是一个由赖氨酸衍生 而成的兼性化合物。它可将脂肪酸以酰基形式从线粒体膜外转运至膜内。其转运机制如下: 肉毒碱与脂酰-CoA 结合生成脂酰肉毒碱,该反应由肉毒碱脂酰转移酶(acyl-CoA transferase)催化,并在线粒体膜外侧进行,脂酰肉毒碱通过线粒体内膜的移位酶 (translocase)穿过内膜,脂酰基与线粒体基质中的辅酶 A 结合,重新产生脂酰-CoA,释