
放肉毒碱。线粒体内膜内侧的肉毒碱转移酶(acyl-CoA transferase)催化此反应。最后经肉 毒碱移位 斯酰CoA 多位 一脂酰碱 骄酰CA 图7-2肉毒碱转运系统 4.脂肪酸B氧化作用的步骤 脂酰C0A进入线粒体后,在基质中进行B氧化作用,包括4个循环步骤: (1)脂酰-CoA的a ,B脱氢作用:脂酰CoA在脂酰-COA脱氢酶 (acyl-CoA dehydrogenase)的催化下,在C和C,(即a、B位)之间脱氢,形成的产物是△2反烯脂 酰-C0A。在线粒体基质中发现有三种脂酰-CoA脱氢酶,分别对短、中、长链的脂肪酸起 专一反应。这3种酶均为黄素蛋白,可与FAD紧密结合,但只催化反式异构体的生成。 脂酰辅牌A脱氢韩 RCH:CH:C~SCoA RCH=CH-( ~SCoA 0 FAD FADHL 0 脂酰CoA △2反烯脂酰-CoA (2)△2-反烯脂酰-CoA的水化:△2-反烯脂酰-CoA在烯脂酰-CoA水合酶(enoyl CoA hydratase)催化下,在双键上加水生成(+B羟脂酰-CoA,此酶具立体化学专一性,只 催化L异构体的生成。 RCH=CH-C一SCOA十H,O,A水合 RCHCH-C~SCoA 0 △2反式烯脂酰:C0A L(+),B-羟脂酰-CoA (3)LB羟脂酰CoA的脱氢作用:在B羟脂酰-CoA脱氢酶(L3-hydr 230
230 放肉毒碱。线粒体内膜内侧的肉毒碱转移酶 (acyl-CoA transferase)催化此反应。最后经肉 毒碱移位酶协助,又回到线粒体外细胞质中(见图 7-2)。 图 7-2 肉毒碱转运系统 4. 脂肪酸β-氧化作用的步骤 脂酰-CoA 进入线粒体后,在基质中进行β-氧化作用,包括 4 个循环步骤: (1)脂酰-CoA 的α、β-脱氢作用:脂酰-CoA 在脂酰-CoA 脱氢酶(acyl-CoA dehydrogenase)的催化下,在 C2 和 C3(即α、β位)之间脱氢,形成的产物是Δ 2反烯脂 酰-CoA。在线粒体基质中发现有三种脂酰-CoA 脱氢酶,分别对短、中、长链的脂肪酸起 专一反应。这 3 种酶均为黄素蛋白,可与 FAD 紧密结合,但只催化反式异构体的生成。 脂酰-CoA Δ2 反烯脂酰-CoA (2)Δ2 -反烯脂酰-CoA 的水化:Δ2 -反烯脂酰-CoA 在烯脂酰-CoA 水合酶(enoyl CoA hydratase)催化下,在双键上加水生成 L(+)-β-羟脂酰-CoA,此酶具立体化学专一性,只 催化 L-异构体的生成。 Δ2 反式烯脂酰-CoA L(+)-β-羟脂酰-CoA (3)L-β-羟脂酰-CoA 的脱氢作用:在β-羟脂酰-CoA 脱氢酶(L-3-hydroxyacyl CoA dehydrogenase )催化下,在 L-β-羟脂酰-CoA 的 C3羟基上脱氢氧化成β-酮脂酰辅酶 A, 反应以 NAD +为辅酶

RHCH,c一sCoA脉满人股室RCCH,一SCoA OH O NAD+ NADH+H+ L(+)-B-羟脂酰.CoA B-酮脂酰-CoA (4)B-酮脂酰-CoA的疏解:在硫解酶(thiolase)即酮脂酰硫解酶催化下B-酮脂酰-CoA 被第二个CoA-SH分子硫解,产生乙酰-CoA和比原来脂酰-CoA少2个碳原子的脂酰 CoA。 g+ RC~SCoA CHC SCoA B酮脂酰辅酶A 脂酰辅酶A 尽管B氧化作用中4个反应步骤都是可逆的,但是由于B酮脂酰CA硫解酶催化的 硫解作用是高度的放能反应(△G,=一28.03km0l),整个反应平衡点趋于裂解方向, 难以进行逆向反应 ,所以使脂肪酸氧化得以继续进行。 经上述5步反应,原脂肪酸脱掉2个碳单位,新形成的脂酰辅酶Λ又可经脱氢、加水、 再脱氢和硫解四步反应进行再一次的B氧化作用。如此重复多次,1分子长链脂肪酸即可 分解成许多分子的乙酰-C0A。B-氧化的整个过程见图7-3。 231
231 L(+)-β-羟脂酰-CoA β-酮脂酰-CoA (4)β-酮脂酰-CoA 的硫解:在硫解酶(thiolase)即酮脂酰硫解酶催化下β-酮脂酰-CoA 被第二个 CoA-SH 分子硫解,产生乙酰-CoA 和比原来脂酰-CoA 少 2 个碳原子的脂酰 -CoA 。 β-酮脂酰辅酶 A 脂酰辅酶 A 尽管β-氧化作用中 4 个反应步骤都是可逆的,但是由于β-酮脂酰-CoA 硫解酶催化的 硫解作用是高度的放能反应(ΔG 0′= -28.03kJ/mol),整个反应平衡点趋于裂解方向, 难以进行逆向反应,所以使脂肪酸氧化得以继续进行。 经上述 5 步反应,原脂肪酸脱掉 2 个碳单位,新形成的脂酰辅酶 A 又可经脱氢、加水、 再脱氢和硫解四步反应进行再一次的β-氧化作用。如此重复多次,1 分子长链脂肪酸即可 分解成许多分子的乙酰-CoA。β-氧化的整个过程见图 7-3
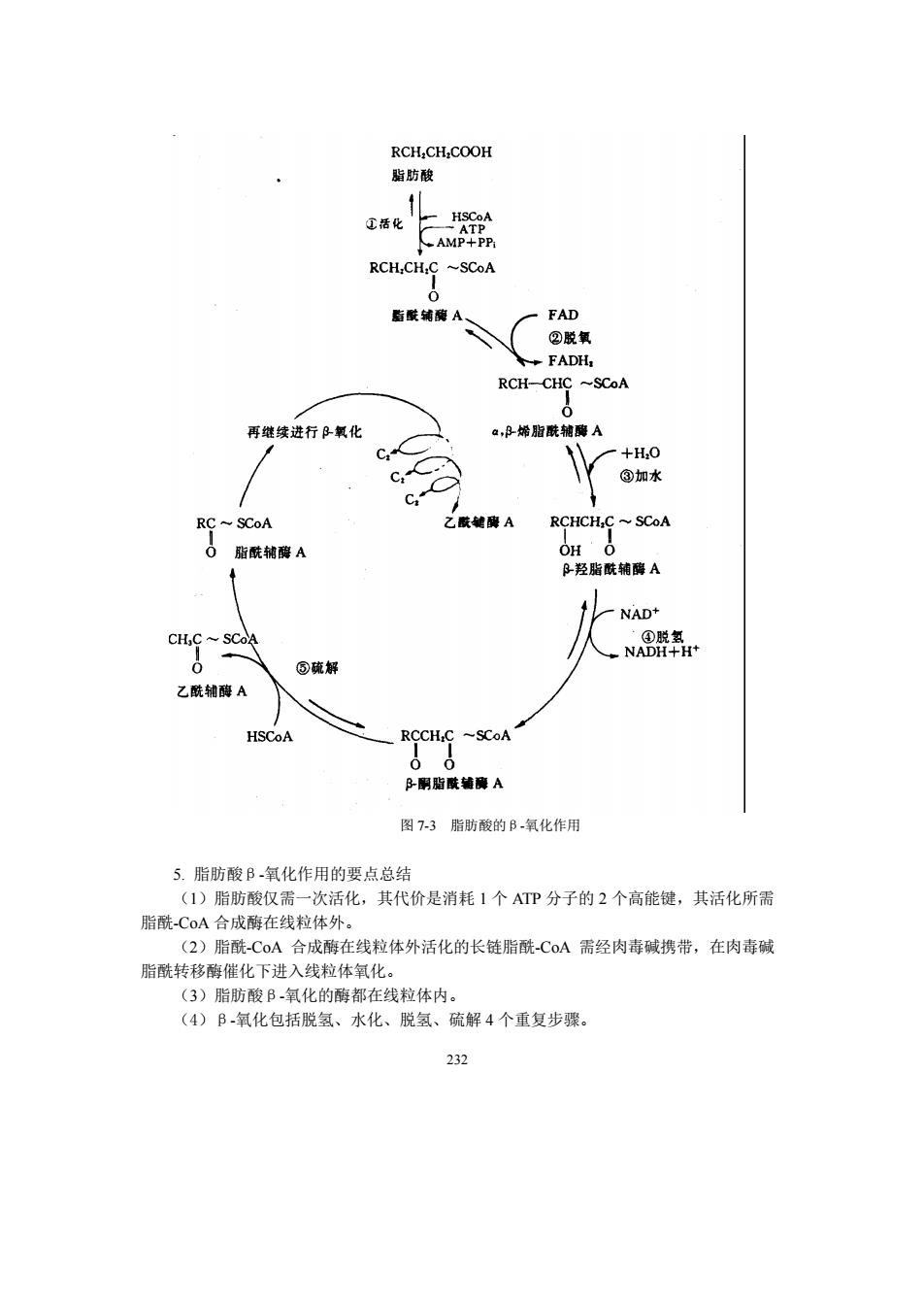
RCH,CH,COOH 酯防酸 ①活化日 AMP+PP RCH.CH.C ~SCoA 后酰铺A、 FAD RCH-CHC~SCoA 再继续进行B氧化 a,B烯脂酰辅德A C +H0 ③加水 RC~SCoA 乙胺能鼻A RCHCH,C~SCoA 0脂酰辅獰A B羟脂酰辅酵A NAD+ CH,C SCoA Na82- @硫解 乙酰辅酶A HSCoA RCCH.C -SCoA B酮贴A 图7-3脂防酸的B氧化作用 5.脂肪酸B氧化作用的要点总结 (1)脂肪酸仅需一次活化,其代价是消耗1个ATP分子的2个高能键,其活化所需 脂酰CoA合成酶在线粒体外。 (2)脂酰C0A合成酶在线粒体外活化的长链脂酰COA需经肉毒碱携带,在肉毒碱 脂酰转移酶催化下进入线粒体氧化, (3)脂肪酸B氧化的酶都在线粒体内。 (4)B氧化包括脱氢、水化、脱氢、硫解4个重复步骤。 232
232 图 7-3 脂肪酸的β-氧化作用 5. 脂肪酸β-氧化作用的要点总结 (1)脂肪酸仅需一次活化,其代价是消耗 1 个 ATP 分子的 2 个高能键,其活化所需 脂酰-CoA 合成酶在线粒体外。 (2)脂酰-CoA 合成酶在线粒体外活化的长链脂酰-CoA 需经肉毒碱携带,在肉毒碱 脂酰转移酶催化下进入线粒体氧化。 (3)脂肪酸β-氧化的酶都在线粒体内。 (4)β-氧化包括脱氢、水化、脱氢、硫解 4 个重复步骤

6.脂防酸B,氧化过程中的能量贮存 脂肪酸B-氧化后形成的乙酰-C0A进入三羧酸循环,最后形成CO2和H,O。 脂肪酸在B氧化中,每形成1分子乙酰-CoA,就使1分子FAD还原为FADH2,并使 I分子NAD还原为NADH+H。FADH进入呼吸链,生成2分子ATP:NADH H进入 呼吸链,生成3分子ATP。因此,每生成1分子乙酰-CoA,就生成5分子ATP。现以软脂 酰-CoA为例,说明其产生ATP分子的过程如下: 软脂酰-COA+HS-COA+FAD+NAD+H,O 豆蔻脂酰CoA+乙酰-COA+FADH,+NADH+H广 经过7次上述的B氧化循环,即可将软脂酰-CoA转变为8分子的乙酰CA 软脂酰-CoA+7HS-CoA+7FAD+7NAD+7HO 8乙酰-CoA+7FADH2+7NADH+7H 每分子乙酰-CoA进入三羧酸循环彻底氧化共生成12分子ATP.因此由8个分子乙酰-COA 氧化为H0和CO2,共形成812=96分子ATP。 由于软脂酸转化为软脂酰-CoA消耗1分子ATP中的两个高能磷酸键的能量,因此净 生成131-2=12 当软脂酸氧化时,自由能的变化是-9790.56 kJ/mol。ATP水解为ADP和Pi时,自由 能的变化为一30.54kJ/mol。软脂酸生物氧化净产生129个ATP,可形成3962.3kJ/mol能量。 因此在软脂酸氧化时约有40%的能量转换成磷酸键能。 (二)脂肪酸的a-氧化 Smp卫K19S6)发现植物线粒体中除有B.氧化作用外,还有1种特殊的氧化途 径,称为a 化作用。这种特殊类型的氧化系统,首先发现于植物种子和 物叶子组织 中,但后来也在脑和肝细胞中发现。在这个系统中,仅游离脂肪酸能作为底物,而且直接 涉及到分子氧,每1次氧化经脂肪酸羧基端只失去1个碳原子,产物既可以是D-ā羟基 脂肪酸,也可以是少1个碳原子的脂肪酸。 脂肪酸α氧化概念是脂肪酸在一些酶的催化下,其ā碳原子发生氧化,结果生成1 分子C0和比原来少1个碳原子的脂肪酸 R-CH2-COOH-R-CH-COOH-R-COOH +CO2 OH ā氧化的机制尚不十分清楚,其可能的途径是: 1.长链脂肪酸在一定条件下,可直接羟化,产生α-羟脂肪酸,再经氧化脱羧作用, 形成以CO,形式去掉一个碳原子的脂肪酸。 233
233 6. 脂肪酸β-氧化过程中的能量贮存 脂肪酸β-氧化后形成的乙酰-CoA 进入三羧酸循环,最后形成 CO2 和 H2O。 脂肪酸在β-氧化中,每形成 1 分子乙酰-CoA,就使 1 分子 FAD 还原为 FADH2,并使 1 分子 NAD +还原为 NADH+H +。FADH2进入呼吸链,生成 2 分子 ATP;NADH+H +进入 呼吸链,生成 3 分子 ATP。因此,每生成 1 分子乙酰-CoA,就生成 5 分子 ATP。现以软脂 酰-CoA 为例,说明其产生 ATP 分子的过程如下: 软脂酰-CoA + HS-CoA + FAD + NAD + + H2O 豆蔻脂酰-CoA +乙酰-CoA + FADH2 + NADH + H + 经过 7 次上述的β-氧化循环,即可将软脂酰-CoA 转变为 8 分子的乙酰-CoA。 软脂酰-CoA + 7HS-CoA + 7FAD + 7NAD + + 7H2O 8 乙酰-CoA + 7FADH2 + 7NADH + 7H + 每分子乙酰-CoA 进入三羧酸循环彻底氧化共生成 12 分子 ATP。因此由 8 个分子乙酰-CoA 氧化为 H2O 和 CO2,共形成 8 12=96 分子 ATP。 由于软脂酸转化为软脂酰-CoA 消耗 1 分子 ATP 中的两个高能磷酸键的能量,因此净 生成 131-2=129 个 ATP。 当软脂酸氧化时,自由能的变化是-9790.56kJ/mol。ATP 水解为 ADP 和 Pi 时,自由 能的变化为-30.54kJ/mol。软脂酸生物氧化净产生 129 个 ATP,可形成 3962.3kJ/mol 能量。 因此在软脂酸氧化时约有 40%的能量转换成磷酸键能。 (二)脂肪酸的α-氧化 Stumpf. P. K.(1956)发现植物线粒体中除有β-氧化作用外,还有 1 种特殊的氧化途 径,称为α-氧化作用。这种特殊类型的氧化系统,首先发现于植物种子和植物叶子组织 中,但后来也在脑和肝细胞中发现。在这个系统中,仅游离脂肪酸能作为底物,而且直接 涉及到分子氧,每 1 次氧化经脂肪酸羧基端只失去 1 个碳原子,产物既可以是 D-α-羟基 脂肪酸,也可以是少 1 个碳原子的脂肪酸。 脂肪酸α-氧化概念是脂肪酸在一些酶的催化下,其α碳原子发生氧化,结果生成 1 分子 CO2 和比原来少 1 个碳原子的脂肪酸。 R CH OH R CH2 COOH COOH R COOH +CO2 α α α-氧化的机制尚不十分清楚,其可能的途径是: 1. 长链脂肪酸在一定条件下,可直接羟化,产生α-羟脂肪酸,再经氧化脱羧作用, 形成以 CO2 形式去掉一个碳原子的脂肪酸
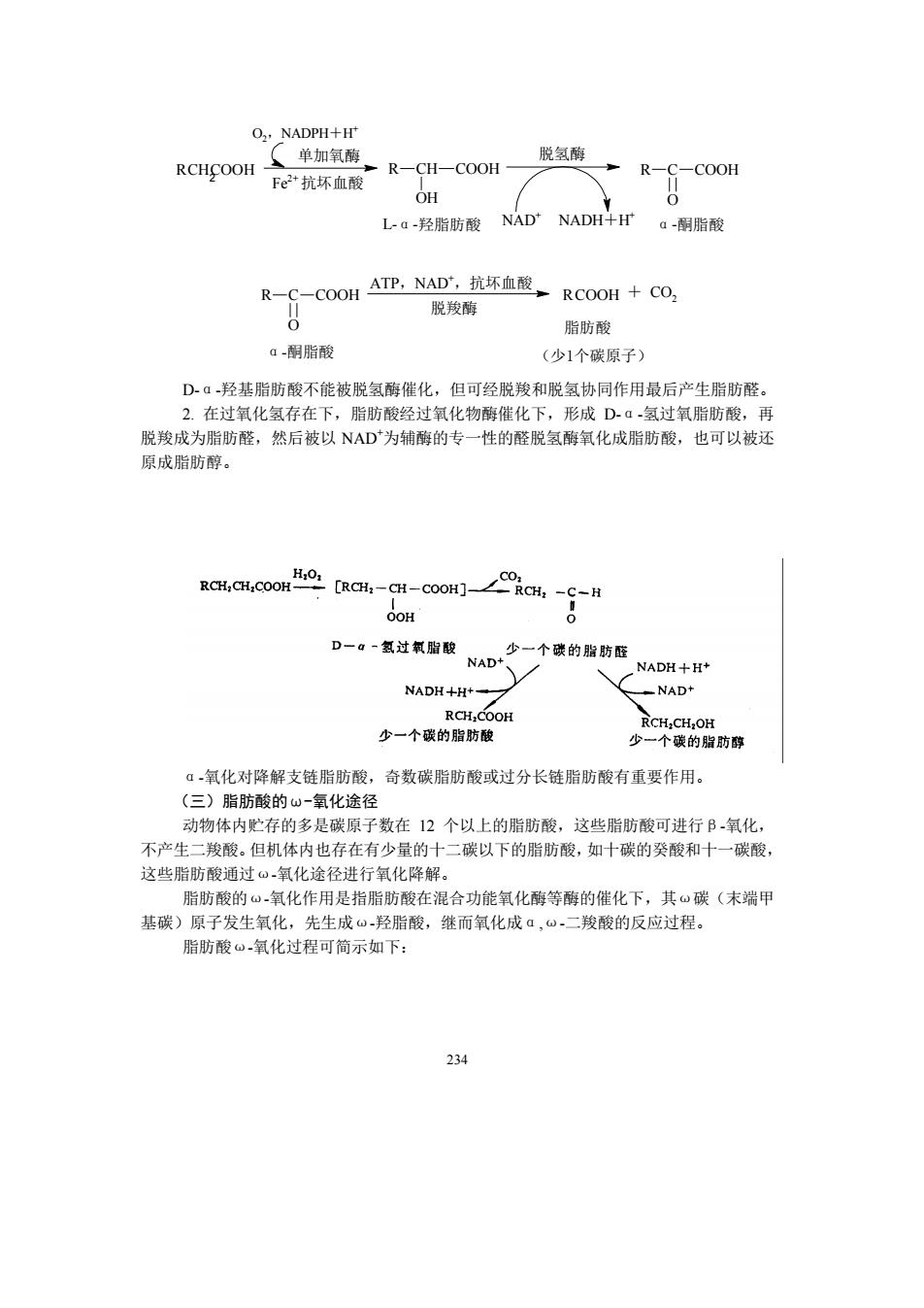
O.NADPH+H' (单加氧酶 R-CH-COOH- 脱氢酶 RCHCOOH Fe2抗坏血酸 →R-C-CoOH OH L-a-羟脂肪酸NAD NADH+Fa-酮脂酸 R-C-C0 OH ATP,NAD抗环直,RC0OH+c0, 脱羧醇 脂肪酸 ā-酮脂酸 (少1个碳原子) Dā羟基胎肪酸不能被脱氢酶催化,但可经脱羧和脱氢协同作用最后产生脂肪醛。 2在过氢化氢存在下,脂肪酸经过氧化物酶催化下,形成D.▣,氢过每脂肪酸。再 脱羧成为脂肪醛,然后被以NAD为辅酶的专一性的醛脱氢酶氧化成脂肪酸,也可以被还 原成脂肪醇。 RCH,CH COO a-o-cwa0∠9a-g OH D一a=复过氧的 NMD个的脂时 .NADH+ NAD+ 少一个碳的脂肪酸 RCH:CH:OH 少一个碳的脂肪醇 ā氧化对降解支链脂肪酸,奇数碳脂肪酸或过分长链胎肪酸有重要作用。 (三)脂肪酸的山-氫化途径 动物体内贮存的多是碳原子数在12个以上的脂肪酸,这些脂肪酸可进行B氧化, 不产生二羧酸。但机体内也存在有少量的十二碳以下的脂肪酸,如十碳的癸酸和十一碳酸, 这些脂肪酸通过氧化途径进行氧化降解。 脂肪酸的“氧化作用是指脂肪酸在混合功能氧化酶等酶的催化下,其~碳(末端甲 基碳)原子发生氧化,先生成⊙羟脂酸,继而氧化成ā,“二羧酸的反应过程。 脂肪酸。氧化过程可简示如下: 234
234 CH2 R COOH O2,NADPH+H + 单加氧酶 Fe 2+抗坏血酸 R CH COOH OH L-α-羟脂肪酸 脱氢酶 NAD + NADH+H + R C COOH O α-酮脂酸 R C COOH O ATP,NAD +,抗坏血酸 脱羧酶 RCOOH + CO2 脂肪酸 α-酮脂酸 (少1个碳原子) D-α-羟基脂肪酸不能被脱氢酶催化,但可经脱羧和脱氢协同作用最后产生脂肪醛。 2. 在过氧化氢存在下,脂肪酸经过氧化物酶催化下,形成 D-α-氢过氧脂肪酸,再 脱羧成为脂肪醛,然后被以 NAD +为辅酶的专一性的醛脱氢酶氧化成脂肪酸,也可以被还 原成脂肪醇。 α-氧化对降解支链脂肪酸,奇数碳脂肪酸或过分长链脂肪酸有重要作用。 (三)脂肪酸的ω-氧化途径 动物体内贮存的多是碳原子数在 12 个以上的脂肪酸,这些脂肪酸可进行β-氧化, 不产生二羧酸。但机体内也存在有少量的十二碳以下的脂肪酸,如十碳的癸酸和十一碳酸, 这些脂肪酸通过ω-氧化途径进行氧化降解。 脂肪酸的ω-氧化作用是指脂肪酸在混合功能氧化酶等酶的催化下,其ω碳(末端甲 基碳)原子发生氧化,先生成ω-羟脂酸,继而氧化成α,ω-二羧酸的反应过程。 脂肪酸ω-氧化过程可简示如下: