
第十章核酸的生物合成 在生物界,物种通过遗传使其生物学特性、形状能世代相传。现代科学已经证明遗 传的物质基础是核酸。核酸是贮存和传递遗传信息的生物大分子。生物体的传信息是 以密码的形式编码在DNA分子上,表现为特定的核苷酸排列顺序。在细胞分裂过程中通 过DNA的复制把遗传信息由亲代传递给子代,在子代的个体发有过程中遗传信息由DNA 传递到RNA,最后翻译成特异的蛋白质,表现出与亲代相似的遗传性状。 1958年,Crik提出了概括遗传信息从一种分子传向另一种分子的理论中心法则 (Central dogma) DA、RNA、蛋白质 这个由DNA决定RNA分子的碱基顺序又由RNA决定蛋白质分子的氨基酸顺序的 理论称为中心法则。 在某些情况下RNA也是重要的遗传物质,如在RNA病毒中,RNA是遗传信息的 带者,具有自我复制的能力,并同时作为mRNA,指导病毒蛋白质的生物合成。在致癌 RNA病毒中,RNA在逆转录酶的作用下,以逆转录(reverse transcription)的方式将遗 传信息RNA传递给DNA分子。 因此,1971年,Cick对中心法则作了进一步补正与完善。修改的中心法则如图10-1 所表示。 复制 毯轻录 DNAY 翻译 蛋白质 图10-1中心法则 图中实线表示遗传信息的一般流向, 虚线表示特殊的流向情况。这个中心法则揭 了核酸与蛋白质合成之间的密切联系和共同规律。 遗传信总的传递和表达主要通过复制、转录和翻译进行。复制(replication)是指以 原来DNA分子为模板,合成出相同DNA分子的过程:转录(transcription)是以DNA 分子为模板合成出与其核苷酸顺序相对应的RNA的过程:翻译(translation)是在由rRNA 和蛋白质组成的核糖核蛋白体(简称核糖体)上 mRNA为 莫板,根据每3个相邻核 苷酸决定一种氨基酸的三联体密码规则,由RNA运送活化的氨基酸,GTP提供所需能 293
293 第十章 核酸的生物合成 在生物界,物种通过遗传使其生物学特性、形状能世代相传。现代科学已经证明遗 传的物质基础是核酸。核酸是贮存和传递遗传信息的生物大分子。生物体的遗传信息是 以密码的形式编码在 DNA 分子上,表现为特定的核苷酸排列顺序。在细胞分裂过程中通 过DNA的复制把遗传信息由亲代传递给子代,在子代的个体发育过程中遗传信息由DNA 传递到 RNA,最后翻译成特异的蛋白质,表现出与亲代相似的遗传性状。 1958 年,Crick 提出了概括遗传信息从一种分子传向另一种分子的理论 中心法则 (Central dogma): DNA RNA 蛋白质 转录 翻译 这个由 DNA 决定 RNA 分子的碱基顺序又由 RNA 决定蛋白质分子的氨基酸顺序的 理论称为中心法则。 在某些情况下 RNA 也是重要的遗传物质,如在 RNA 病毒中,RNA 是遗传信息的携 带者,具有自我复制的能力,并同时作为 mRNA,指导病毒蛋白质的生物合成。在致癌 RNA 病毒中,RNA 在逆转录酶的作用下,以逆转录(reverse transcription)的方式将遗 传信息 RNA 传递给 DNA 分子。 因此,1971 年,Crick 对中心法则作了进一步补正与完善。修改的中心法则如图 10-1 所表示。 RNA 蛋白质 翻 译 D N A 复 制 逆 转 录 转 录 复制 图 10-1 中心法则 图中实线表示遗传信息的一般流向,虚线表示特殊的流向情况。这个中心法则揭示 了核酸与蛋白质合成之间的密切联系和共同规律。 遗传信息的传递和表达主要通过复制、转录和翻译进行。复制(replication)是指以 原来 DNA 分子为模板,合成出相同 DNA 分子的过程;转录(transcription)是以 DNA 分子为模板合成出与其核苷酸顺序相对应的 RNA 的过程;翻译(translation)是在由 rRNA 和蛋白质组成的核糖核蛋白体(简称核糖体)上,以 mRNA 为模板,根据每 3 个相邻核 苷酸决定一种氨基酸的三联体密码规则,由 tRNA 运送活化的氨基酸,GTP 提供所需能

量,合成出具有特定氨基酸顺序的蛋白质肽链的过程 第一节DNA的生物合成 DNA分子是由两条多核苷酸链组成的,它可以在生物体内进行合成作用。合成作用 进行时,需要有一条模板链指导多核苷酸链合成中核苷酸的排列顺序。以DNA作为模板 指导的DNA合成作用,复制出新的DNA分子,如此将DNA携带的信总传递给子代DNA DNA的合成也能以RNA为模板,反转录合成DNA分子,这种反转录合成常见于RNA 病毒。环境因素可以造成DNA结构发生损伤,损伤的DNA可进行修复((repair)合成, 即校正错误的序列,继续进行正确的合成反应,以保持遗传信息的稳定性。 一、半保留复制 DNA呈双股螺旋结构,这样的结构对于维持遗传物质的稳定性和复制的准确性都是 极为重要的。两条链是严格遵循AT和 C碱基配X 新形成的氢健联合在一起,这两条 链是互补的。一条链上的核苷酸顺序决定另一条链核苷酸排列顺序。Watson和crick提出 DNA双螺旋结构模型时推测,在DNA复制时,亲代DNA的双螺旋先行解旋和分开,然 后以每条链为模板,按照碱基配对原则,在这两条链上各形成一条互补链(图10-2)。这 样,从亲代DNA的分子可以精确地复制成2个子代DNA分子。每个子代DNA分子中, 有一条链是从亲代 DNA来的,另一条则是新形成的,这叫做 半保留复制 replication)(图10- 与此时也有人提 全保留复制方式,即复制后亲代DNA的两 条链不变,子代DNA的两条链都是新合成的。I957年Meselson及Stahl通过实验证实了 半保留复制的模式。 294
294 量,合成出具有特定氨基酸顺序的蛋白质肽链的过程。 第一节 DNA 的生物合成 DNA 分子是由两条多核苷酸链组成的,它可以在生物体内进行合成作用。合成作用 进行时,需要有一条模板链指导多核苷酸链合成中核苷酸的排列顺序。以 DNA 作为模板 指导的DNA合成作用,复制出新的DNA分子,如此将DNA携带的信息传递给子代DNA。 DNA 的合成也能以 RNA 为模板,反转录合成 DNA 分子,这种反转录合成常见于 RNA 病毒。环境因素可以造成 DNA 结构发生损伤,损伤的 DNA 可进行修复(repair)合成, 即校正错误的序列,继续进行正确的合成反应,以保持遗传信息的稳定性。 一、半保留复制 DNA 呈双股螺旋结构,这样的结构对于维持遗传物质的稳定性和复制的准确性都是 极为重要的。两条链是严格遵循 A-T 和 G-C 碱基配对所形成的氢键联合在一起,这两条 链是互补的。一条链上的核苷酸顺序决定另一条链核苷酸排列顺序。Watson 和 crick 提出 DNA 双螺旋结构模型时推测,在 DNA 复制时,亲代 DNA 的双螺旋先行解旋和分开,然 后以每条链为模板,按照碱基配对原则,在这两条链上各形成一条互补链(图 10-2)。这 样,从亲代 DNA 的分子可以精确地复制成 2 个子代 DNA 分子。每个子代 DNA 分子中, 有一条链是从亲代 DNA 来的,另一条则是新形成的,这叫做半保留复制(semiconservative replication)(图 10-3)。与此同时也有人提出全保留复制方式,即复制后亲代 DNA 的两 条链不变,子代 DNA 的两条链都是新合成的。1957 年 Meselson 及 Stahl 通过实验证实了 半保留复制的模式

、亲代DNA分子 T -A -A- 4.C G- G.:-CS c-G SA-T A C- T G G ∠C-G TA- A -TA A C.S ∠G -T-A 复制桂 G 亲本链复制链 T·-A 亲本链 图1O-2胎tson和Cmck提出的DNA双螺旋复制模型 以大肠杆菌作为实验材料,在培养基中生长繁殖。首先在培养基中以1N标记的 NH,C1作为氨的唯一来源(即重培养基)。大肠杆菌在重培养基中繁殖约15代(每代约 20~30min),DNA可全部为lN所标记,然后将细菌转移到含有N标记NH,CI的培养 基(即轻培养基)中进行培养。在培养不同代数时,收集细菌,裂解细胞,用CsC1密度 梯度离心法分析DNA。实验结果表明,在重培养基中培养出的(N)DNA显示为一条 重密度带。转入轻培养基中繁殖两代。第一代得到了一条中密度带,这是由于其为(NN) 295
295 图 10-2 Watson 和 Crick 提出的 DNA 双螺旋复制模型 以大肠杆菌作为实验材料,在培养基中生长繁殖。首先在培养基中以 15N 标记的 NH4Cl 作为氮的唯一来源(即重培养基)。大肠杆菌在重培养基中繁殖约 15 代(每代约 20~30min),DNA 可全部为 15N 所标记,然后将细菌转移到含有 14N 标记 NH4Cl 的培养 基(即轻培养基)中进行培养。在培养不同代数时,收集细菌,裂解细胞,用 CsCl 密度 梯度离心法分析 DNA。实验结果表明,在重培养基中培养出的(15N)DNA 显示为一条 重密度带。转入轻培养基中繁殖两代。第一代得到了一条中密度带,这是由于其为(15N 14N)
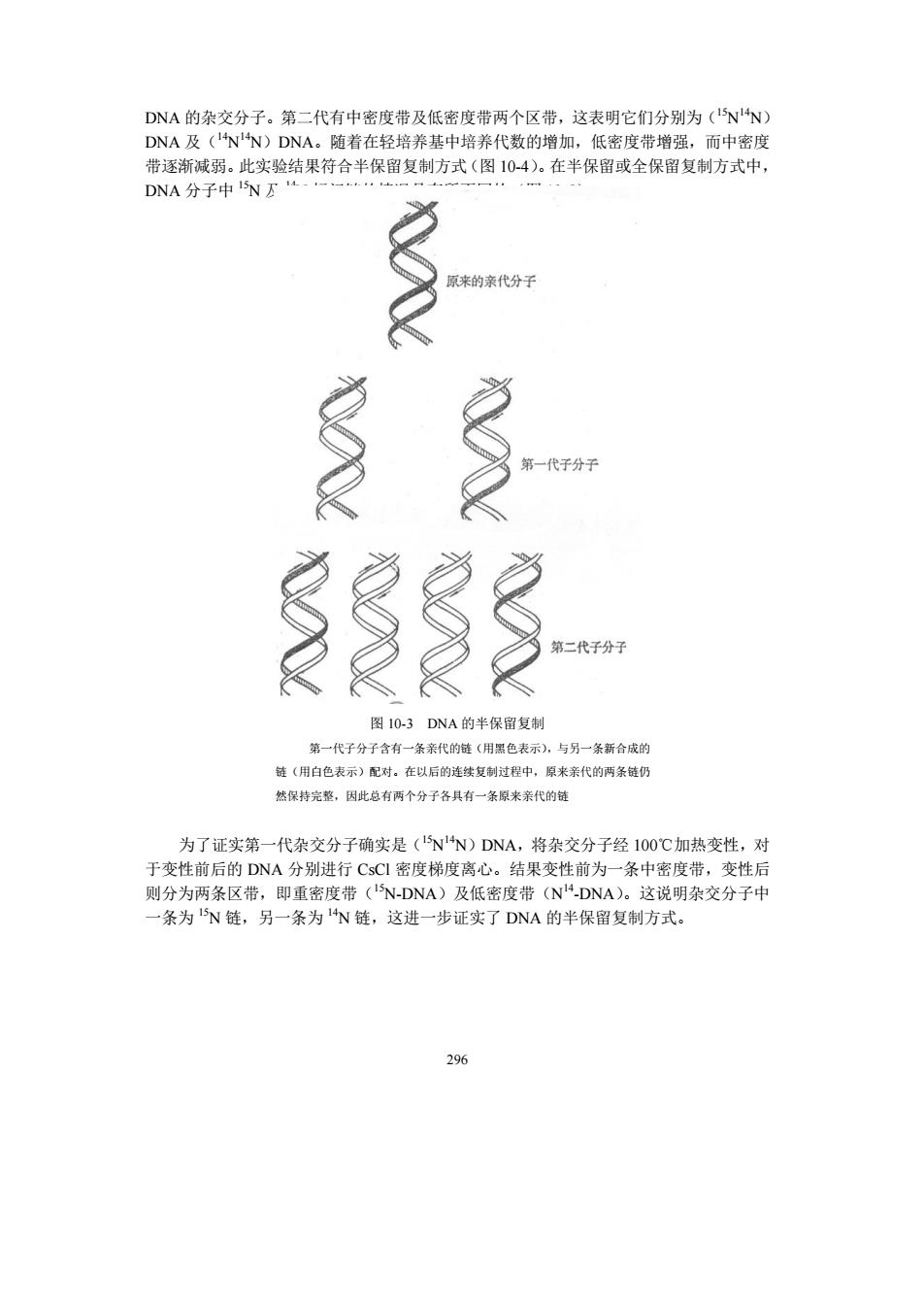
DNA的杂交分子。第二代有中密度带及低密度带两个区带,这表明它们分别为(NN) DNA及(NN)DNA。随着在轻培养基中培养代数的增加,低密度带增强,而中密度 带逐渐减弱。此实验结果符合半保留复制方式(图104)。在半保留或全保留复制方式中, DNA分子中NE 原来的亲代分子 多 二代子分子 图I0-3DNA的半保留复制 第一代子分子含有一条亲代的鼓《用黑色表示).与另一条新合成 链《用白色表示)配对。在以后的续复制过程中,原米亲代的两条链仍 然保持完整,因此总有两个分子各具有一条原来来代的链 为了证实第一代杂交分子确实是(NN)DNA,将杂交分子经1O0C加热变性,对 于变性前后的DNA分别进行CsCI密度梯度离心。结果变性前为一条中密度带,变性后 则分为两条区带,即重密度带(NDNA)及低密度带(NDNA)。这说明杂交分子中 条为N链,另一条为N链,这进一步证实了DNA的半保留复制方式。 296
296 DNA 的杂交分子。第二代有中密度带及低密度带两个区带,这表明它们分别为(15N 14N) DNA 及(14N 14N)DNA。随着在轻培养基中培养代数的增加,低密度带增强,而中密度 带逐渐减弱。此实验结果符合半保留复制方式(图 10-4)。在半保留或全保留复制方式中, DNA 分子中 15N 及 14N 标记链的情况是有所不同的(图 10-5)。 图 10-3 DNA 的半保留复制 第一代子分子含有一条亲代的链(用黑色表示),与另一条新合成的 链(用白色表示)配对。在以后的连续复制过程中,原来亲代的两条链仍 然保持完整,因此总有两个分子各具有一条原来亲代的链 为了证实第一代杂交分子确实是(15N 14N)DNA,将杂交分子经 100℃加热变性,对 于变性前后的 DNA 分别进行 CsCl 密度梯度离心。结果变性前为一条中密度带,变性后 则分为两条区带,即重密度带(15N-DNA)及低密度带(N 14 -DNA)。这说明杂交分子中 一条为 15N 链,另一条为 14N 链,这进一步证实了 DNA 的半保留复制方式

亲代分子 第一代子代分于 0 第二代子代分 1.0 M 19 轻密度DNA 图104N标记的DNA在N培养基中复制两代时的实验结果 (a)DNA区智的紫外吸收图 (b)光密度 N 在培养蒸中复制 亲代 在、培养燕中复制! 子代 半保留复制 全保留复制 图I0-5半保留及全保留复制时DNA中标记链的分布 297
297 图 10-4 15N 标记的 DNA 在 14N 培养基中复制两代时的实验结果 (a)DNA 区带的紫外吸收图 (b)光密度圈 15N 15N 15 N 15N 在 15N 培养基中复制 亲代 在 15N 培养基中复制 ↓ ↓ 15N 14N 14N 15N 14N 14N 15N 15N 子代 半保留复制 全保留复制 图 10-5 半保留及全保留复制时 DNA 中标记链的分布