
第十一章蛋白质的生物合成 蛋白质是生命活动的重要物质基础,要不断地进行代谢和更新,因此,细胞内利用20 种基本氨基酸进行蛋白质合成便成了生命现象的主要内容。以大肠杆菌为例,蛋白质占细 胞的干重50%左右,每个细胞约有3000种不同的蛋白质分子,每种蛋白质又有无数分子。 而大肠杆菌细胞的分裂周期不过20mi,可见蛋白质生物合成的速度之快。 目前已经完全清楚,细胞内每个蛋白质分子的生物合成都受到细胞内DW的指导,但 是贮存遗传信总的DNA并非蛋白质合成的直接模板(template)。它是经转录作用把遗传信 息传递到信使核糖核酸(messenger ribonucleic acid.mRNA)的结构中,所以mRNA才是 蛋白质合成的直接模板。mRNA是由4种核苷酸构成的多核苷酸,而蛋白质是由20种左 右的氨基酸构成的多肽,它们之间遗传信息的传递并不像转录那么简单。从多核苷酸上所 携带的遗传信息,到多肽链上所携带的遗传信息的传递,与从一种语言翻译成另一种语言 时的情形相似。所以人们称以mRNA为模板的蛋白质合成过程为翻译或转译(translation)。 可以将生物遗传信息的传递归纳为以下图解(图1山1): 转录 DNA RNA译、蛋白质 反转录 复制(病毒) 图11!生物遗传信息的传递方向 翻译的过程十分复杂,几乎涉及到细胞内所有种类的NA和几十种蛋白质因子,其中 包括有核糖体RNA(ribosomal ribonucleic acid,rRNA)、信使RNA、转移RNA(transfer ,RNA)、氨酰-tRNA合成酶(aminoacyl-iRNA synthetase)以及一些辅助因 子,即起始因子(initiation factor.F)、延伸因子(elongation factor.EF)、释放因子(终l 因子)(release factor,RF)等参加的协同作用。 在蛋白质合成中,RNA按mRNA模板的要求将相应的氨基酸搬运到蛋白质合成的场 所 核糖体(ribosome)上,所以把核糖体称作蛋白质合成的工厂,氨基酸之间以肽键连 接,生成具有一定排列顺序的蛋白质。蛋白质合成的原料是氨基酸,反应所需能量由ATP 和GTP提供。 蛋白质生物合成的早期研究工作都是用原核生物(prokarvotes)大肠杆菌的无细胞体系 327
327 第十一章 蛋白质的生物合成 蛋白质是生命活动的重要物质基础,要不断地进行代谢和更新,因此,细胞内利用 20 种基本氨基酸进行蛋白质合成便成了生命现象的主要内容。以大肠杆菌为例,蛋白质占细 胞的干重 50%左右,每个细胞约有 3 000 种不同的蛋白质分子,每种蛋白质又有无数分子。 而大肠杆菌细胞的分裂周期不过 20min,可见蛋白质生物合成的速度之快。 目前已经完全清楚,细胞内每个蛋白质分子的生物合成都受到细胞内 DNA的指导,但 是贮存遗传信息的 DNA 并非蛋白质合成的直接模板(template)。它是经转录作用把遗传信 息传递到信使核糖核酸(messenger ribonucleic acid, mRNA)的结构中,所以 mRNA 才是 蛋白质合成的直接模板。mRNA 是由 4 种核苷酸构成的多核苷酸,而蛋白质是由 20 种左 右的氨基酸构成的多肽,它们之间遗传信息的传递并不像转录那么简单。从多核苷酸上所 携带的遗传信息,到多肽链上所携带的遗传信息的传递,与从一种语言翻译成另一种语言 时的情形相似。所以人们称以 mRNA 为模板的蛋白质合成过程为翻译或转译(translation)。 可以将生物遗传信息的传递归纳为以下图解(图 11-1): 图 11-1 生物遗传信息的传递方向 翻译的过程十分复杂,几乎涉及到细胞内所有种类的 RNA和几十种蛋白质因子,其中 包括有核糖体 RNA(ribosomal ribonucleic acid, rRNA)、信使 RNA、转移 RNA(transfer ribonucleic acid, tRNA)、氨酰-tRNA合成酶(aminoacyl-tRNA synthetase)以及一些辅助因 子,即起始因子(initiation factor, IF)、延伸因子(elongation factor, EF)、释放因子(终止 因子)(release factor, RF)等参加的协同作用。 在蛋白质合成中,tRNA 按 mRNA 模板的要求将相应的氨基酸搬运到蛋白质合成的场 所 核糖体(ribosome)上,所以把核糖体称作蛋白质合成的工厂,氨基酸之间以肽键连 接,生成具有一定排列顺序的蛋白质。蛋白质合成的原料是氨基酸,反应所需能量由 ATP 和 GTP 提供。 蛋白质生物合成的早期研究工作都是用原核生物(prokaryotes)大肠杆菌的无细胞体系 蛋白质 翻 译 复 制 D N A 复制(病毒) 转 录 反转录 R N A

(ce-free system)进行的。所以,对大肠杆菌的蛋白质合成机理了解最多,真核生物 (cukaryotes).的蛋白质合成的机理与大肠杆菌的有许多相似之处,但也有不少差异,下面即 详细讨论原核生物与真核生物的蛋白质的生物合成过程。 第一节蛋白质合成体系的重要组分 一、mRNA与遗传密码 (一)信使RNA概念的提出 信使RNA(messenger RNA,mRNA)的发现在分子生物学的发展中是一重大事件。由于 其在细胞,总RNA中所占比例很小,很难把它分离出来。mRNA的概念首先是从理论上提 出来的,然后再用实验得到证实。F.Jacob和J.Monod早在1961年就提出mRNA的概念。 他们认为,既然蛋白质是在胞质中合成的,而编码蛋白质的信息载体DNA却在胞核内 那么必定有一种中间物质用来传递DNA上的信总。他们在研究大肠杆菌中与乳糖代谢有 关醇类的生物合成时发现,诱导物如异丙基硫代半乳糖苷(B., PTG)的加入, 可以立刻使酶蛋白的合成速度增加上千倍。而诱导物 旦消失,又可使酶 蛋白的合成立刻停止。这个实验结果给人的启示是:蛋白质合成的模板是一种不稳定的物 质,其半衰期很短。他们对这种信使物质的性质作了如下的预言: 1.信使是一种多核苷酸: 2.信使的碱基组成应与相应的DNA的碱基组成相一致: 3.信使的长度应是不同的,因为由它们所编码的多肽链的长度是不同的 4.在多肽合成时信使应与核糖体作短暂的结合: 5信使的半期很短,所以信使的合成速度应该是很快的。 所以,这样的信使可能是一种RNA。但是当时已发现的两种RNA(RNA、RNA 都不具备这些特性。各种生物的核糖体RNA的大小差异不大,碱基组成的变化也不大。 RNA除了有与RNA相同的问题以外,它们的分子也太小。所以这两种RNA都不能胜任 信使的功能。可喜的是当时已有人提出过,细胞内有可能存在第三种RNA。在被噬菌 T,感染后的大肠杆菌中,有人发现有一种新的NA,它的代谢速度极快,分子大小也参 差不齐,碱基组成又与TDNA相一致。这些特征都符合信使分子的要求 (仁)信使RNA的实验证明 信使RNA的概念提出后,还必须要用实验来证明这种概念是否正确。为此,S.Brenner E.Jacob和4.Monocl等人设计了一组实验。用噬菌体T,感染大肠杆菌后,发现几乎所有 在细胞内合成的蛋白质都不再是细胞本身的蛋白质,而是噬菌体所编码的蛋白质:这些蛋 328
328 (cell-free system)进行的。所以,对大肠杆菌的蛋白质合成机理了解最多,真核生物 (eukaryotes)的蛋白质合成的机理与大肠杆菌的有许多相似之处,但也有不少差异,下面即 详细讨论原核生物与真核生物的蛋白质的生物合成过程。 第一节 蛋白质合成体系的重要组分 一、mRNA 与遗传密码 (一)信使 RNA概念的提出 信使 RNA(messenger RNA, mRNA)的发现在分子生物学的发展中是一重大事件。由于 其在细胞总 RNA 中所占比例很小,很难把它分离出来。mRNA 的概念首先是从理论上提 出来的,然后再用实验得到证实。F. Jacob 和 J. Monod 早在 1961 年就提出 mRNA 的概念。 他们认为,既然蛋白质是在胞质中合成的,而编码蛋白质的信息载体 DNA 却在胞核内, 那么必定有一种中间物质用来传递 DNA 上的信息。他们在研究大肠杆菌中与乳糖代谢有 关酶类的生物合成时发现,诱导物如异丙基硫代半乳糖苷(β-isopropylthiogalaotoside, IPTG)的加入,可以立刻使酶蛋白的合成速度增加上千倍。而诱导物一旦消失,又可使酶 蛋白的合成立刻停止。这个实验结果给人的启示是:蛋白质合成的模板是一种不稳定的物 质,其半衰期很短。他们对这种信使物质的性质作了如下的预言: 1.信使是一种多核苷酸; 2.信使的碱基组成应与相应的 DNA 的碱基组成相一致; 3.信使的长度应是不同的,因为由它们所编码的多肽链的长度是不同的; 4.在多肽合成时信使应与核糖体作短暂的结合; 5.信使的半衰期很短,所以信使的合成速度应该是很快的。 所以,这样的信使可能是一种 RNA。但是当时已发现的两种 RNA(rRNA、tRNA) 都不具备这些特性。各种生物的核糖体 RNA 的大小差异不大,碱基组成的变化也不大。 tRNA 除了有与 rRNA 相同的问题以外,它们的分子也太小。所以这两种 RNA 都不能胜任 信使的功能。可喜的是当时已有人提出过,细胞内有可能存在第三种 RNA。在被噬菌体 T2 感染后的大肠杆菌中,有人发现有一种新的 RNA,它的代谢速度极快,分子大小也参 差不齐,碱基组成又与 T2DNA 相一致。这些特征都符合信使分子的要求。 (二)信使 RNA的实验证明 信使 RNA 的概念提出后,还必须要用实验来证明这种概念是否正确。为此,S.Brenner, F. Jacob 和 M. Monocl 等人设计了一组实验。用噬菌体 T2感染大肠杆菌后,发现几乎所有 在细胞内合成的蛋白质都不再是细胞本身的蛋白质,而是噬菌体所编码的蛋白质;这些蛋
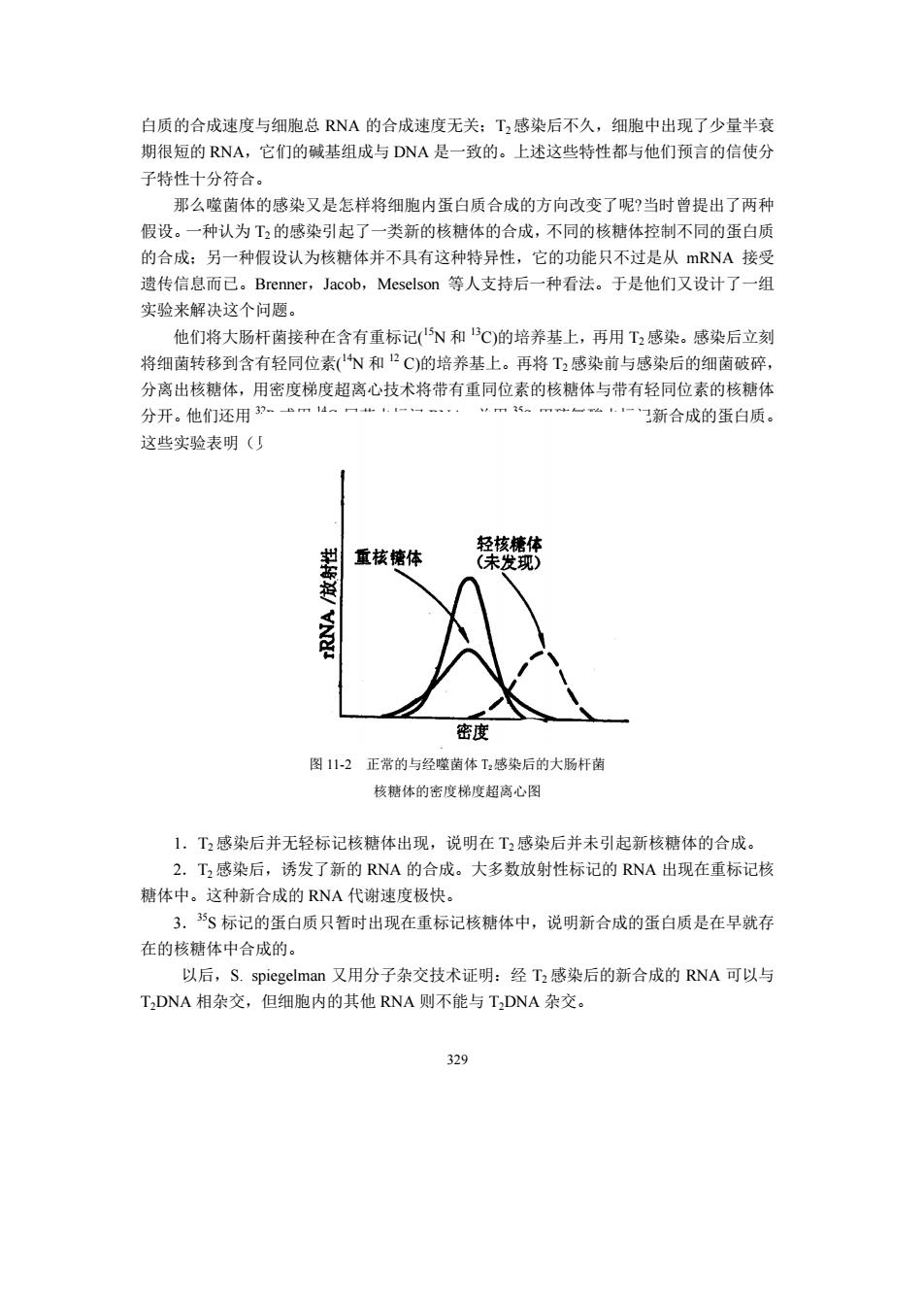
白质的合成速度与细胞总RNA的合成速度无关:T2感染后不久,细胞中出现了少量半衰 期很短的RNA,它们的碱基组成与DNA是一致的。上述这些特性都与他们预言的信使分 子特性十分符合。 那么噬菌体的感染又是怎样将细胞内蛋白质合成的方向改变了呢?当时曾提出了两种 假设。一种认为T2的感染引起了一类新的核糖体的合成,不同的核糖体控制不同的蛋白质 的合成:另一种假设认为核糖体并不具有这种特异性,它的功能只不过是从mRNA接受 遗传信息而己。Brenner,Jacob,Meselson等人支持后一种看法。于是他们又设计了一组 实验来解决这个问题。 他1将大肠杆南接种在含有重标记N和C)的培养基上,再用T,成染。感染后立刻 将细菌转移到含有轻同位素N和PC)的培养基上。再将感染前与感染后的细菌破碎 分离出核糖体,用密度梯度超离心技术将带有重同位素的核糖体与带有轻同位素的核糖体 分开。他们还用m4一“一一 一一一门新合成的蛋白质。 这些实验表明() 割 密度 图12正常的与经嘴菌体T,感染后的大肠杆菌 核糖体的密度梯度超离心图 1.工感染后并无轻标记核糖体出现,说明在T2感染后并未引起新核糖体的合成。 2.T,成染后,诱发了新的RNA的合成。大名数放射性标记的RNA出现在重标记核 糖体中。这种新合成的RNA代谢速度极快。 3.5、标记的蛋白质只暂时出现在重标记核糖体中,说明新合成的蛋白质是在早就存 在的核糖体中合成的。 以后,S.spiegelman又用分子杂交技术证明:经T2感染后的新合成的RNA可以与 T,DNA相杂交,但细胞内的其他RNA则不能与TDNA杂交。 329
329 白质的合成速度与细胞总 RNA 的合成速度无关;T2感染后不久,细胞中出现了少量半衰 期很短的 RNA,它们的碱基组成与 DNA 是一致的。上述这些特性都与他们预言的信使分 子特性十分符合。 那么噬菌体的感染又是怎样将细胞内蛋白质合成的方向改变了呢?当时曾提出了两种 假设。一种认为 T2的感染引起了一类新的核糖体的合成,不同的核糖体控制不同的蛋白质 的合成;另一种假设认为核糖体并不具有这种特异性,它的功能只不过是从 mRNA 接受 遗传信息而已。Brenner,Jacob,Meselson 等人支持后一种看法。于是他们又设计了一组 实验来解决这个问题。 他们将大肠杆菌接种在含有重标记( 15N 和 13C)的培养基上,再用 T2 感染。感染后立刻 将细菌转移到含有轻同位素( 14N 和 12 C)的培养基上。再将 T2 感染前与感染后的细菌破碎, 分离出核糖体,用密度梯度超离心技术将带有重同位素的核糖体与带有轻同位素的核糖体 分开。他们还用 32 P 或用 14C-尿苷去标记 RNA,并用 35 S-甲硫氨酸去标记新合成的蛋白质。 这些实验表明(见图 11-2): 图 11-2 正常的与经噬菌体 T2感染后的大肠杆菌 核糖体的密度梯度超离心图 1.T2 感染后并无轻标记核糖体出现,说明在 T2 感染后并未引起新核糖体的合成。 2.T2 感染后,诱发了新的 RNA 的合成。大多数放射性标记的 RNA 出现在重标记核 糖体中。这种新合成的 RNA 代谢速度极快。 3.35 S 标记的蛋白质只暂时出现在重标记核糖体中,说明新合成的蛋白质是在早就存 在的核糖体中合成的。 以后,S. spiegelman 又用分子杂交技术证明:经 T2 感染后的新合成的 RNA 可以与 T2DNA 相杂交,但细胞内的其他 RNA 则不能与 T2DNA 杂交

(三)责传密码 1.遗传密码的发现 mRNA是蛋白质合成的直接模板,其核苷酸排列顺序取决于相应DNA的碱基排列顺 序,它又决定了所形成的蛋白质多肽链中的氨基酸的排列顺序。那么mRNA上的核苷酸 排列顺序是如何翻译成蛋白质中的氨基酸的排列顺序,即如何编码成遗传密码的呢? mRNA中有4种核苷酸,用数学方法推算,如果每一种核苷酸代表一种氨基酸,那么 只能代表4种氨基酸。如果每两个相邻的核苷酸代表一种氨基酸,可以有4-16种排列方 式,显然也不足以代表20种基本氨基酸。如果每三个相邻的核苷酸代表一种氨基酸,可 以有4-64种排列方式,这就足以满足为20种基本氨基酸编码的需要。所以这种编码方 式的可能性最大。应用生物化学和遗传学研究技术,已经证明是三个相邻的核苷酸编码 种氨基酸,这三个连续的核苷酸称为三联体密码((riplet code)或密码子(codon).。 如何证明密码子和氨基酸之间的对应关系?1961年,Nirenberg等用大肠杆菌无细胞体 系,外加20种标记氨基酸混合物及0vU,经保温反应后,发现在酸不溶性部分中(即多 肽中)只有苯丙氨酸的多聚体。所以UUU是编码苯丙氨酸的密码。同样,用polyA和pOlC 作为mRNA来合成蛋白质,结果分别只得到多聚赖氨酸和多聚脯氨酸,说明AAA是赖氨 酸的密码,CCC是脯氨酸的密码。 进一步,Nirenberg和Ochoa等用polyUG,polyAC重复上述类似实验,发现标记氨基 酸掺入新合成的肽链的频率与按统计学方法推算出的多核苷酸中三联体密码出现的频率 相符合。即 ol(UG):UGU GUG UGU GUG UGU GUG UGU GUG 翻译成:Cvs-Val-Cvs.Val-Cvs-Val poly(AC):ACACACACA CAC ACACACACACAC 翻译成:Thr-His-Thr-His-Thr-His 应用这种方法,仅用了四年时间,于1965年完全查清了20种基本氨基酸所对应的全 部61个密码子,其余三个密码子为终止密码子,编出了遗传密码字典(表111)。 330
330 (三)遗传密码 1.遗传密码的发现 mRNA 是蛋白质合成的直接模板,其核苷酸排列顺序取决于相应 DNA 的碱基排列顺 序,它又决定了所形成的蛋白质多肽链中的氨基酸的排列顺序。那么 mRNA 上的核苷酸 排列顺序是如何翻译成蛋白质中的氨基酸的排列顺序,即如何编码成遗传密码的呢? mRNA 中有 4 种核苷酸,用数学方法推算,如果每一种核苷酸代表一种氨基酸,那么 只能代表 4 种氨基酸。如果每两个相邻的核苷酸代表一种氨基酸,可以有 4 2 =16 种排列方 式,显然也不足以代表 20 种基本氨基酸。如果每三个相邻的核苷酸代表一种氨基酸,可 以有 4 3 =64 种排列方式,这就足以满足为 20 种基本氨基酸编码的需要。所以这种编码方 式的可能性最大。应用生物化学和遗传学研究技术,已经证明是三个相邻的核苷酸编码一 种氨基酸,这三个连续的核苷酸称为三联体密码(triplet code)或密码子(codon)。 如何证明密码子和氨基酸之间的对应关系?1961 年,Nirenberg 等用大肠杆菌无细胞体 系,外加 20 种标记氨基酸混合物及 polyU,经保温反应后,发现在酸不溶性部分中(即多 肽中)只有苯丙氨酸的多聚体。所以 UUU是编码苯丙氨酸的密码。同样,用 polyA 和 polyC 作为 mRNA 来合成蛋白质,结果分别只得到多聚赖氨酸和多聚脯氨酸,说明 AAA 是赖氨 酸的密码,CCC 是脯氨酸的密码。 进一步,Nirenberg 和 Ochoa 等用 polyUG,polyAC 重复上述类似实验,发现标记氨基 酸掺入新合成的肽链的频率与按统计学方法推算出的多核苷酸中三联体密码出现的频率 相符合。即 poly(UG): UGU GUG UGU GUG UGU GUG UGU GUG 翻译成: Cys-Val-Cys-Val-Cys-Val poly(AC): ACA CAC ACA CAC ACA CAC ACA CAC 翻译成: Thr-His-Thr-His-Thr-His 应用这种方法,仅用了四年时间,于 1965 年完全查清了 20 种基本氨基酸所对应的全 部 61 个密码子,其余三个密码子为终止密码子,编出了遗传密码字典(表 11-1)
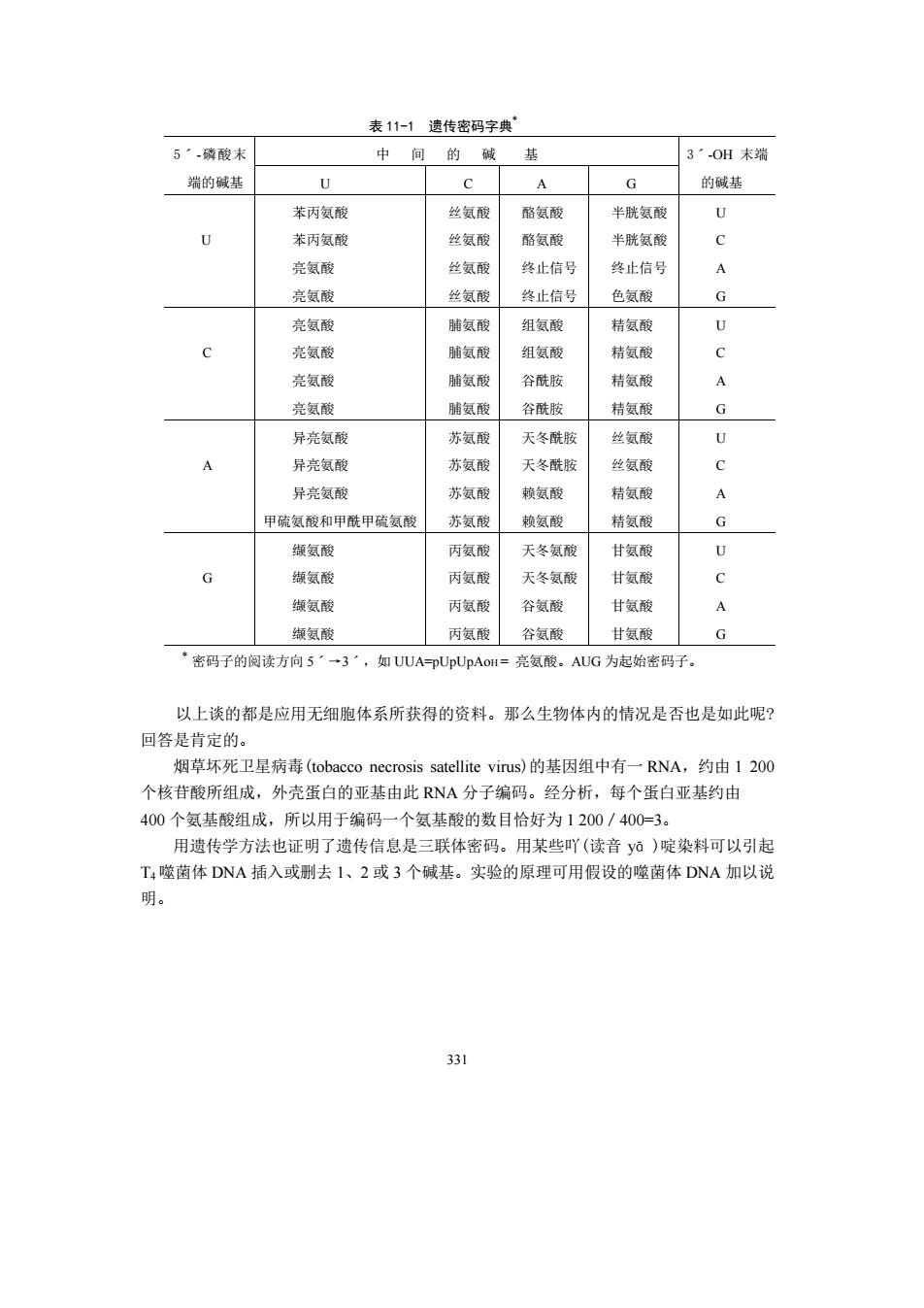
表11-1遗传密码字典 5·-磷酸末 中间的碱 基 3'-OH末端 端的碱基 A 的碱基 苯丙氨酸 丝氨酸 酪氨酸 半胱氨酸 U U 苯丙氨酸 丝氨酸 酪氨酸 半胱氨酸 c 亮氨解 掉氨的 终止信号 终止信号 A 亮氨酯 丝氨酸 终止信号 色氨酸 G 亮氨酸 躺氨酸 组氨酸 精氨酸 亮氨酪 氨酸 组氨酸 精氨酶 c 亮氨酸 脂氨酸 谷酰胺 精氨酸 A 亮氨酸 氨酸 谷酰胺 精氨酸 G 异亮氨酸 苏氨酸 天冬酰胺 丝氨 U 异亮氨酸 苏氨酸 天冬酰酸 丝氨酾 异亮氨酸 苏氨酸 氨酸 精氨酸 甲硫氨酸和甲酰甲硫氨酸 苏氨酸 赖氨酸 精氨酸 G 缬氨酸 丙氨酸 天冬氨酸 甘氨酸 11 缬氨酸 丙氨剥 天冬氨酸 甘氨酸 C 缬氨酸 丙氨酸 谷氨酸 甘氨酸 A 数氨酸 丙氨酸谷氨酸 甘氨酸 G ‘密码子的阅读方向5'一3',如UUA-pUpUpAot=亮氨酸。AUG为起始密码子. 以上谈的都是应用无细胞体系所获得的资料。那么生物体内的情况是否也是如此呢? 回答是肯定的。 烟草坏死卫星病毒(tobacco necrosis satellite virus)的基因组中有一RNA,约由1200 个核苷酸所组成,外壳蛋白的亚基由此RNA分子编码。经分析,每个蛋白亚基约由 400个氨基酸组成,所以用于编码一个氨基酸的数目恰好为1200/400=3。 用遗传学方法也证明了遗传信息是三联体密码。用某些吖(读音yā)啶染料可以引起 T噬菌体DNA插入或去1、2或3个碱基。实验的原理可用假设的噬茵体DNA加以说 明。 331
331 表 11-1 遗传密码字典 * 5ˊ-磷酸末 中 间 的 碱 基 端的碱基 U C A G 3ˊ-OH 末端 的碱基 U 苯丙氨酸 苯丙氨酸 亮氨酸 亮氨酸 丝氨酸 丝氨酸 丝氨酸 丝氨酸 酪氨酸 酪氨酸 终止信号 终止信号 半胱氨酸 半胱氨酸 终止信号 色氨酸 U C A G C 亮氨酸 亮氨酸 亮氨酸 亮氨酸 脯氨酸 脯氨酸 脯氨酸 脯氨酸 组氨酸 组氨酸 谷酰胺 谷酰胺 精氨酸 精氨酸 精氨酸 精氨酸 U C A G A 异亮氨酸 异亮氨酸 异亮氨酸 甲硫氨酸和甲酰甲硫氨酸 苏氨酸 苏氨酸 苏氨酸 苏氨酸 天冬酰胺 天冬酰胺 赖氨酸 赖氨酸 丝氨酸 丝氨酸 精氨酸 精氨酸 U C A G G 缬氨酸 缬氨酸 缬氨酸 缬氨酸 丙氨酸 丙氨酸 丙氨酸 丙氨酸 天冬氨酸 天冬氨酸 谷氨酸 谷氨酸 甘氨酸 甘氨酸 甘氨酸 甘氨酸 U C A G *密码子的阅读方向 5ˊ→3ˊ,如 UUA=pUpUpAoH = 亮氨酸。AUG 为起始密码子。 以上谈的都是应用无细胞体系所获得的资料。那么生物体内的情况是否也是如此呢? 回答是肯定的。 烟草坏死卫星病毒(tobacco necrosis satellite virus)的基因组中有一 RNA,约由 1 200 个核苷酸所组成,外壳蛋白的亚基由此 RNA 分子编码。经分析,每个蛋白亚基约由 400 个氨基酸组成,所以用于编码一个氨基酸的数目恰好为 1 200/400=3。 用遗传学方法也证明了遗传信息是三联体密码。用某些吖(读音 yā)啶染料可以引起 T4 噬菌体 DNA 插入或删去 1、2 或 3 个碱基。实验的原理可用假设的噬菌体 DNA 加以说 明